生命理工学系 News
シクリッドゲノム中に適応進化の痕跡を発見
祖先から受け継いだゲノム多様性が急速な進化の鍵
要点
- 進化のモデル生物・シクリッド3種18個体の全ゲノム配列を解読
- 適応進化の候補遺伝子の多くに祖先多型が存在していることを明らかに
- シクリッドの急速な適応進化には祖先多型が寄与した可能性を示唆
概要
東京工業大学 生命理工学院 生命理工学系の二階堂雅人准教授(生命理工学コース主担当)、中村遥奈大学院生、相原光人研究員、伊藤武彦教授(生命理工学コース主担当)、梶谷嶺助教(生命理工学コース主担当)、岡田典弘名誉教授およびタンザニア水産研究所、国立遺伝学研究所、北里大学の共同研究チームは、進化研究のモデル生物と称されるシクリッドの全ゲノム配列の解析を通じて、急速な適応進化には祖先から受け継いだゲノムの多様性(祖先多型[用語1] )が重要な役割を果たしたことを明らかにした。
本研究では短期間に適応進化を遂げたビクトリア湖産シクリッドの中から、特に3種に着目してその全ゲノム配列を決定し(各種6個体/計18個体)、それらを網羅的に集団遺伝解析[用語2] することで、シクリッドの適応進化に関わったと考えられる遺伝子を数多く単離することに成功した。そして、さらなる詳細な系統解析により、これら適応遺伝子には古い祖先種から受け継がれてきたゲノムの多様性が、大規模な「祖先多型」として存在することが明らかになった。
一般に生物のゲノム中において、祖先多型は時間と共に失われてしまうと考えられており、シクリッドが進化の過程で失わずに受け継いできた、この例外的に大規模な祖先多型こそが、適応放散[用語3] を可能にしたゲノム基盤であることを示唆している。
この研究成果は3月22日(日本時間)に米国の学術誌『Molecular Biology and Evolution』電子版にて公開された。
背景
東アフリカの大地溝帯(アフリカ大陸を南北に縦断する巨大な谷)に位置する三大湖(ビクトリア湖、マラウィ湖、タンガニィカ湖)には、形態的・生態的に多様なシクリッド(淡水性の熱帯魚)が数百種を超えて生息しており、これらは各湖に固有の種であることが分かっている(図1)。
また、これらのシクリッドは各湖において急速な適応放散を遂げたことが知られており、形態や生態が如何にして進化・多様化するのかを理解するためのモデル生物として世界の研究者が注目している。
特にビクトリア湖産シクリッドは、適応放散を遂げてからわずか15,000年しか経過していない事が地質学的な研究から示唆されているため、この極めて短い期間で蓄積し得るごく僅かなDNAの違いによって、驚くべき多様性が生み出されたと予想されてきた。
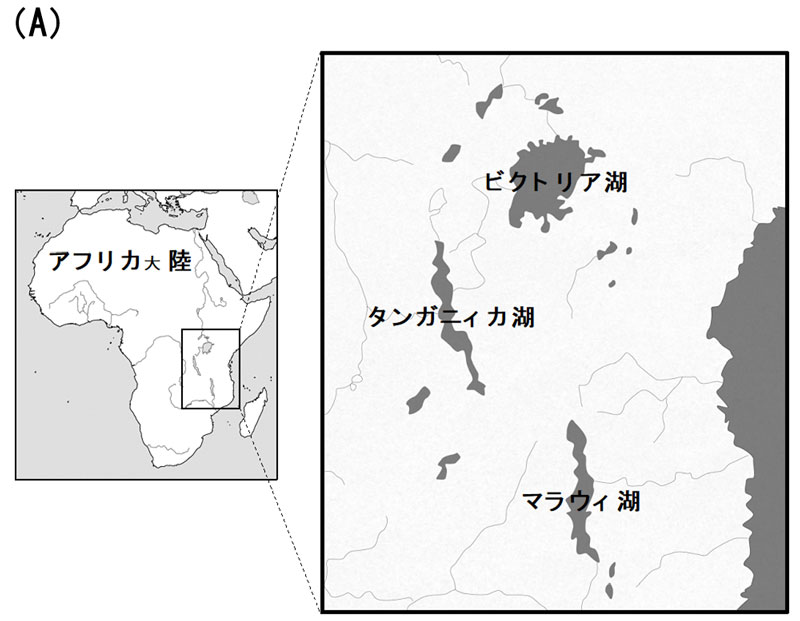

図1.(A)東アフリカ三大湖と(B)本研究でゲノム解読を実施したビクトリア湖産シクリッド。(A)タンガニィカ湖とマラウィ湖は大地溝帯の裂け目に存在し、ビクトリア湖はその間に挟まれて存在。(B)上からLithochromis rufus, Haplochromis sauvagei, H. chilotes。この3種は15,000年前に共通祖先から適応放散によって分化してきた極めて近縁な種にもかかわらず、体サイズ、体色や形態だけでなくその生息環境も多様である。
この東アフリカ産シクリッドの適応進化に関わる遺伝的基盤を明らかにする目的で、国際研究チームは2014年に三大湖各1種と周辺河川2種のシクリッド計5種について全ゲノム配列を決定した(注1)。この解析ではシクリッドのゲノム中に正の自然淘汰[用語4] の痕跡を数多く発見することに成功しただけでなく、それまでシクリッドの進化を議論する中で注目されることのなかった「祖先多型」の存在も明らかにし、シクリッドの適応進化研究の礎を築いた。
その後も東アフリカ産シクリッドの多くの種を網羅したゲノム解析が続けられ、このグループの進化の全貌がゲノムレベルで少しずつ明らかになりつつある。しかし、シクリッドの適応進化を遺伝子レベルで明らかにするためには、1種について1個体ずつゲノム配列を解読するだけでは不十分であった。つまり、各種について複数個体のゲノム配列を網羅的に解析することにより、種間で分化したゲノム領域を特定する必要があった。
(注1)2014年10月21日 東工大ニュース「アフリカ・シクリッドの多様性は過去のゲノム多型が基盤 ―シクリッド5種の全ゲノム配列を決定して解明―」
研究成果
本研究では種の適応進化に関わる遺伝子を探索する目的で、生態や形態の異なるビクトリア湖産シクリッド3種(Haplochromis sauvagei, H. chilotes, Lithochromis rufus:図1B参照)に着目して、それぞれの種について6個体(計18個体)の全ゲノム配列を決定し、それらを集団遺伝学的な手法を用いて解析した。
我々がもっとも重要な研究目的として掲げていたのが、適応に関わる遺伝子やその遺伝的変異の特定であった。今回の研究では、種間における分化の程度を示す値Fst[用語5] に着目して、このFstの高い領域を探索した。ビクトリア湖産シクリッドは適応放散から15,000年しか経過していないため、種間分化の程度も低く、実際にゲノム全体のFstは平均すると0.1~0.2程度であった。
それに対し、適応進化に関わり種間で異なる生態や形態を生み出すゲノム領域は、自然選択によって種間で分化し、Fstの値も高くなると予想される。そこで今回はFstの値が0.58~0.75以上(上位0.5%)のゲノム領域を、適応や分化に関わる候補とした。
また今回の研究ではFstに加えて、塩基配列の違いの指標となるDxy[用語6] を推定し、FstとDxyが共に高い領域を探索することで、近縁な種であるにもかかわらず、高度に分化した領域を積極的に探索した。その結果、行動や免疫機構などに関わる計726個の遺伝子を、特に種間で高度に分化している遺伝子として単離することに成功した。(図2:コラーゲン遺伝子領域の例)
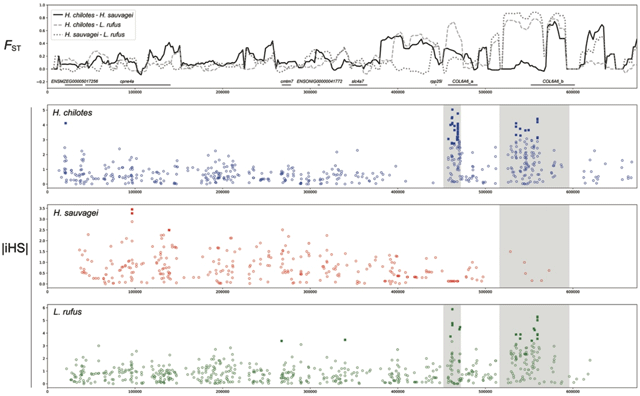
図2.種間で高度に分化した適応候補遺伝子の一例
コラーゲン6A6遺伝子の分化した2つのアリルはマラウィ湖やタンガニィカ湖のシクリッドのゲノム中にも存在しており、これが祖先多型に由来したことが明らかになった。コラーゲン6A6遺伝子は、Fstの値だけでなくiHSの値でも種間で高度に分化した遺伝子であることが示された。iHSは各集団において特定領域での自然選択の痕跡を検出する指標の一つである。今回の研究では、コラーゲン6A6遺伝子領域に強い正の自然選択が働いた可能性が示唆された。
今回の研究結果で特筆すべき点として、上記の適応進化の候補遺伝子は、種間で分化していると同時に、多くの塩基置換[用語7] が蓄積していることが挙げられる。常に一定とされる突然変異の速度から考えると、ビクトリア湖の適応放散から15,000年でこれほどの塩基置換が蓄積するとは考えにくい。我々は、この遺伝子がどのような過程を経ることで、予想を超えた塩基置換を蓄積するに至ったのかを明らかにするため、これら適応候補遺伝子の系統解析をおこなった。
その結果、適応候補遺伝子の多くについて、ビクトリア湖のシクリッド種間で分化したアリル[用語8] のどちらもが、マラウィ湖やタンガニィカ湖のシクリッドにも存在している事が明らかとなった。これは三大湖のシクリッドの共通祖先の段階ですでに存在していた遺伝子の多様性が、シクリッドの進化の過程で失われずに維持されたまま、祖先多型として各湖に再分配されたことを意味している(図3)。
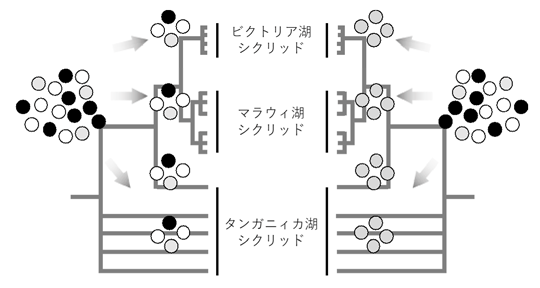
図3.東アフリカ産シクリッドの系統樹と祖先多型の分配(モデル)
今回の研究で、ビクトリア湖のシクリッド種間で分化した複数のアリルが、マラウィ湖やタンガニィカ湖のシクリッドにも共有されていたことから、その多様性は東アフリカ産シクリッドの共通祖先までさかのぼることが示唆された(左)。一般的には、複数存在するアリルは時間の経過と共にどれか1つに固定すると予想されるが(右)、シクリッドではそれらのアリル多様性が何らかの機構で祖先多型として維持されたと考えられる。この多様なアリルに自然選択が働くことで、急速な適応が可能になったと予想される。色のついた丸印は集団中に存在する異なるアリルを示す。
種内における遺伝子の多様性は一定の時間の経過と共に徐々に失われていくことがわかっており、一般的な進化パターンを考えると、今回のように大規模な祖先多型が維持されることは論理的には考えにくい。東アフリカ産シクリッドの進化過程における交雑や集団サイズの増大、地理的なイベントなどいくつかの要因が重なることで、大規模な祖先多型が維持されたと予想される。そして、適応放散の前から祖先多型としてすでに存在していた遺伝的多様性(多様なアリル)に自然選択が働くことで、シクリッドの急速な適応進化が可能になったのだと考えられる。
今後の展開
今回の研究でシクリッドのゲノム中に発見された大規模な祖先多型が、進化の過程でどのようにして維持されてきたのか、そして、祖先多型がゲノムの中でも特に適応進化に関わる領域に偏って存在するメカニズムを明らかにするために、シミュレーション等を駆使した詳細な検証研究を進める予定である。また、祖先多型に由来する適応進化の候補遺伝子が、実際にシクリッドの種間における形態や生態の分化と結びついているのかどうかを、遺伝子改変シクリッドを用いた分子生物学的な機能解析によって実証していく予定である。
研究グループ
本研究は、東京工業大学 生命理工学院 生命理工学系の二階堂雅人准教授、中村遥奈大学院生(日本学術振興会特別研究員DC2)、相原光人研究員、伊藤武彦教授、梶谷嶺助教、タンザニア水産研究所のHillary D. J. Mrosso(ヒラリー・ムロッソ)博士、Semvua I. Mzighani(センブア・ムジガニ)博士、国立遺伝学研究所の豊田敦特任教授、北里大学の岡田典弘特任教授(東工大名誉教授)によって進められた。
- 用語説明
[用語1] 祖先多型 : 集団を構成する個体間でのDNA配列の違いを多型という。多型は時間と共に失われてしまうため、集団(種)形成後に突然変異や交雑により獲得された多型が集団内の遺伝的多様性の多くを占める。一方で、祖先多型は集団(種)形成より前の祖先集団において獲得され、何らかの機構により維持されている多型を指す。
[用語2] 集団遺伝解析 : 集団(種)内の複数個体のゲノムデータからその遺伝的多様性に着目し、生物集団の進化過程を解析する方法。例えば、個体数の増減等の過去の歴史の推定や集団間比較による自然淘汰を受けた遺伝子の同定を行なうことができる。
[用語3] 適応放散 : 祖先となる少数の種が生息範囲を広げ異なる環境に適応することで、短期間に形態学・生態学的に多様な種が爆発的に生じること。
[用語4] 正の自然淘汰 : 有利な変異がその適応度の高さから集団内に広がり、最終的には固定する過程のこと。集団が生息する環境において有利な変異を持つ個体は持たない個体に比べ、適応度が上がり、次世代に子孫を多く残す。その結果、有利な変異は集団内で多くの割合を占めるようになり、最終的には固定する。
[用語5] Fst : 集団間の塩基配列の違いの相対的指標の一つ。2集団間の遺伝的分化の程度を各集団の遺伝的多様性を考慮して定量化している。0~1の値をとり、遺伝的分化の程度が大きいほど、大きな値をとる。
[用語6] Dxy : 集団間の塩基配列の違いの絶対的指標の一つ。遺伝的分化の程度を2集団間の塩基配列の違いから計算している。遺伝的分化の程度が大きいほど、大きな値をとる。
[用語7] 塩基置換 : DNAにおいてA(アデニン)、T(チミン)、G(グアニン)、C(シトシン)の4種類の塩基の並びを塩基配列と呼び、遺伝情報が内包されている。塩基置換は配列内のある塩基が複製の間違いなどにより別の塩基に置き換わることを指す。
[用語8] アリル : 同一のゲノム領域(遺伝子座)に存在する、両親から受け継いだそれぞれの遺伝子のことを指す。対立遺伝子とも呼ぶ。
- 論文情報
| 掲載誌 : | Molecular Biology and Evolution |
|---|---|
| 論文タイトル : | Genomic Signatures for Species-Specific Adaptation in Lake Victoria Cichlids Derived from Large-Scale Standing Genetic Variation |
| 著者 : | Haruna Nakamura, Mitsuto Aibara, Rei Kajitani, Hillary D. J. Mrosso, Semvua I. Mzighani, Atsushi Toyoda, Takehiko Itoh, Norihiro Okada, Masato Nikaido |
| DOI : |
- カモノハシとハリモグラの全ゲノム解読に成功!|生命理工学系 News
- オオコウモリ2種の全ゲノム配列を解読|生命理工学系 News
- フェロモン受容機構が退化した哺乳類をゲノム解析で特定|生命理工学系 News
- ほぼ全ての脊椎動物に共通するフェロモン受容体を発見|生命理工学系 News
- アフリカ・シクリッドの多様性は過去のゲノム多型が基盤 ―シクリッド5種の全ゲノム配列を決定して解明―|東工大ニュース
- タンザニア北部にシーラカンスの繁殖集団を発見|東工大ニュース
- 【研究室紹介】二階堂研究室 ―生物多様性創出の分子メカニズムの解明―|生命理工学系 News
- 二階堂研究室
- 研究者詳細情報(STAR Search) - 二階堂雅人 Masato Nikaido
- 研究者詳細情報(STAR Search) - 相原光人 Mitsuto Aibara
- 研究者詳細情報(STAR Search) - 伊藤武彦 Takehiko Itoh
- 研究者詳細情報(STAR Search) - 梶谷嶺 Rei Kajitani
- 研究者詳細情報(STAR Search) - 岡田典弘 Norihiro Okada
- 生命理工学院 生命理工学系
- 国立遺伝学研究所
- 北里大学
- 研究成果一覧
お問い合わせ先
東京工業大学 生命理工学院 生命理工学系
准教授 二階堂雅人
E-mail : mnikaido@bio.titech.ac.jp
Tel : 03-5734-2659 / Fax : 03-5734-2946
おすすめ記事
-

2026.07.24
-

2026.07.23
-

2026.07.22
-

2026.07.22
-
2026.07.17
![]()