生命理工学系 News
光合成機能の制御が生産性維持に重要であることを解明
植物の光合成酵素の制御スイッチを壊したら何が起こるのか
要点
- ゲノム編集技術で植物の光合成酵素の機能制御スイッチを壊した
- 酵素そのものは常に活性化状態になった
- 通常の光条件では問題なく成長したが、変動光では成長が阻害された
概要
東京工業大学 生命理工学院 生命理工学系 横地佑一大学院生(研究当時)と科学技術創成研究院 化学生命科学研究所の久堀徹教授(ライフエンジニアリングコース主担当)らは、ドイツのハインリッヒ・ハイネ大学のアンドレアス・ウエバー教授らと共同で、植物の光合成を制御するレドックス制御機構[用語1]の生理的な役割を調べるため、光合成酵素のひとつNADP-リンゴ酸脱水素酵素(MDH)[用語2]の制御スイッチをゲノム編集技術で破壊し、この制御機構の植物における重要性を明らかにした。
光合成を行う葉緑体には、変化する自然環境に応じて光合成に関わる酵素の活性を調節する「レドックス(酸化還元)スイッチ」が備わっている。葉緑体の代謝機能は環境変化によって変動するため、葉緑体内のいろいろな酵素の活性を変化に応じて調節することが、機能の維持に不可欠と考えられてきた。
そこで「効率的な光合成には酵素活性の調節が重要」との仮説のもと葉緑体のレドックス調節に関する研究が行われてきたが、光合成を営む植物が生きていく上で調節機構がどんな役割を果たすのかは明らかになっていなかった。
この仮説を検証するため、研究グループはMDHのレドックススイッチを壊した場合の植物の代謝への影響を調べた。ゲノム編集技術でMDH が2つ持っているレドックススイッチの1つをなくし、暗所でも活性がオフにならない酵素を持つ変異植物を作製した。この変異植物は暗所でもMDHの働きでNADPH[用語3]を消費するため生育に不利と予想されたが、生育実験に用いられる標準的な長日条件では野生型の植物と大きな違いが見られなかった。ところが、短日条件や明暗が頻繁に切り替わる変動光条件では変異型の植物は明らかに生育が遅延した。すなわち、制御スイッチによるMDH 活性の制御は、常に周囲の状況が変動する自然環境のような条件下では、葉緑体内の還元物質NADPHの量をあるレベルに保って代謝機能を維持するために重要であるということである。
この研究成果は2月3日(日本時間)に「Proceedings of National Academy of Science USA」電子版に発表された。
背景
植物の光合成は、光エネルギーを用いて炭水化物を生産する、地球規模の壮大なエネルギー変換反応である。光合成を営む植物の緑葉の細胞は、葉緑体という細胞内小器官を多数持っており、ここで光合成反応を行っている。個々の葉緑体の内部には、チラコイド膜とよばれる生体膜が積層した形で発達している。
光合成電子伝達系[用語4]を構成する膜タンパク質は、このチラコイド膜に埋めこまれており、ここで光エネルギーを捕獲して化学エネルギー(ATPとNADPH)への変換反応が行われている。光合成電子伝達系で生産されたATPとNADPHは、葉緑体のストロマとよばれる膜以外のゾル部分に存在する酵素群の働きによって、大気中の二酸化炭素から炭水化物を生産する化学反応に用いられる。
この二酸化炭素の固定反応は、発見者の名前を取ってCalvin-Benson(カルビン・ベンソン)回路とよばれている。Calvin-Benson回路は、昼間、電子伝達系が光エネルギーを捕捉するときに働くように制御されており、この制御機構はレドックス制御と呼ばれる。この制御機構は、光合成の電子伝達系で生じた還元力の一部が還元物質であるNADPHの生成に使われず、葉緑体内で働く酵素分子が持っているジスルフィド結合[用語5]の還元に用いられることによって、個々の酵素の活性が制御される、というものである。
Calvin-Benson回路では、グリセルアルデヒド3-リン酸デヒドロゲナーゼ(GAPDH)、フルクトース1,6ビスフォスファターゼ(FBPase)、セドヘプツロース1,7ビスフォスファターゼ(SBPase)、ホスホリブロキナーゼ(PRK)などが、このようなレドックス制御を受ける酵素として以前から知られている。葉緑体内では、これらの酵素以外にもATP合成酵素や、リンゴ酸脱水素酵素(MDH)がレドックス制御を受けている。さらに、2000年以降のプロテオミクス技術[用語6]の進展で、数多くの酵素がこのような制御を受けていることが明らかになってきた。
このように、葉緑体内でいろいろな酵素がレドックス制御を受けている理由は、光合成が行える環境下でのみ効率よく酵素反応を進行させるためと考えられてきた。酵素は化学反応の触媒なので、熱力学的にはどちらの方向にも反応を進めることができる。簡単に言えば、糖を合成する酵素は条件さえ整えば糖を分解することもできるということになる。
光合成の条件下では、光エネルギーを化学エネルギーに変換する方向ですべての反応が進行して、葉緑体の中では糖が合成される。光が当たらないときには、酵素によっては逆反応を触媒して糖の分解方向に反応を進行させることがあっても不思議はない。しかし、明るい所で合成した糖を暗い所で分解していたら、昼間と夜が交互にやってくる環境に住む植物にとっては極めて不都合である。そこで、葉緑体のレドックス制御システムは、夜間に働くと不都合な酵素の活性をレドックス制御によってオフにしていると解釈されている。
ところが、これまでレドックス制御が本当に植物の生存に必須の機能であることを明確に示した生理学的な実験というのはほとんど行われていない。たとえば、レドックス制御の鍵タンパク質としていろいろな酵素に還元力を伝達するチオレドキシンというタンパク質がある。植物細胞内に複数種あるチオレドキシンを正常に機能させないと植物の生育に影響することが知られている。しかし、この実験は、昼間オンになっている必要がある酵素の機能を停止すると生育が影響されるということを示しているだけで、レドックス制御の重要な働きと考えられる“夜間の酵素活性をオフにする”必要が本当にあるのかはこれまでわかっていなかった。
葉緑体のMDHは、葉緑体内でNADPHとオキサロ酢酸からリンゴ酸とNADP+を生成する酵素である。NADPHは生体膜を通過することができないが、リンゴ酸は葉緑体の外に容易に出ることができる。還元物質であるNADPHは、葉緑体内ではCalvin-Benson回路で糖の合成に使われている。さらに、葉緑体内のNADPHの還元力は、MDHによってリンゴ酸に渡され葉緑体の外にも輸送されるわけだ。葉緑体の外に出たリンゴ酸を用いて細胞質のMDHがNADHのような還元物質を生成し、これが他の代謝系で使われる。このような還元力の輸送システムをリンゴ酸バルブと呼ぶ(図1)。
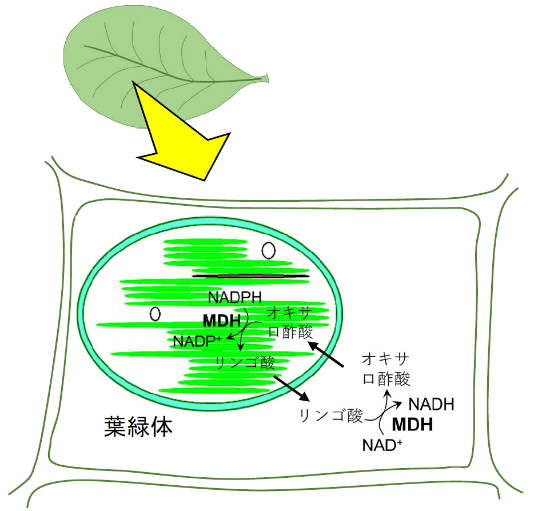
図1. 葉緑体のリンゴ酸バルブ
もし、光合成を行わない夜間にも葉緑体のMDHが働くと、リンゴ酸バルブが還元力を輸送し続けて葉緑体内の還元力が枯渇してしまうと考えられる。そこで、葉緑体のMDHでは、レドックス制御によって夜間には不都合な活性がオフにされていると予想されていた。研究チームは、このMDHのレドックス制御スイッチを働かなくしたら何が起こるのかを調べることによって、レドックス制御の生理的な役割を明らかにすることにした。
研究成果
MDHという酵素は、原核生物から真核生物まで広く生物界に存在し、還元物質の変換という重要な役割を担っている。葉緑体のMDHと細菌や他の生物のMDHのアミノ酸配列を比較すると、葉緑体型のMDHだけがN末端側とC末端側[用語7]に伸長した配列を持っており、なおかつ、この伸長部分にそれぞれレドックス制御スイッチとして働く1組のCys[用語8]を持っている(図2A)。つまり、葉緑体MDH分子には、2つのレドックス制御スイッチが備えられているわけだ。
研究チームは、まず葉緑体MDHのC末端側のアミノ酸配列を書き換え、C末端側を部分的に欠損していて2個目のレドックス制御スイッチが壊れている組換え体タンパク質[用語9]を大腸菌を用いて作成した(図2B)。
野生型タンパク質も変異型タンパク質も完全に還元すると高い活性を示した(図2C、四角)。逆に酸化してジスルフィド結合を分子内に作ると野生型の酵素の活性はほとんどゼロになった(図2C、黒三角)。ところが変異型タンパク質は、活性がちょうど野生型の酸化状態と還元状態の中間くらいになった(図2C、赤三角)。すなわち、C末端側のレドックススイッチが壊れると、酸化されても酵素が完全には止まらなくなるわけだ。
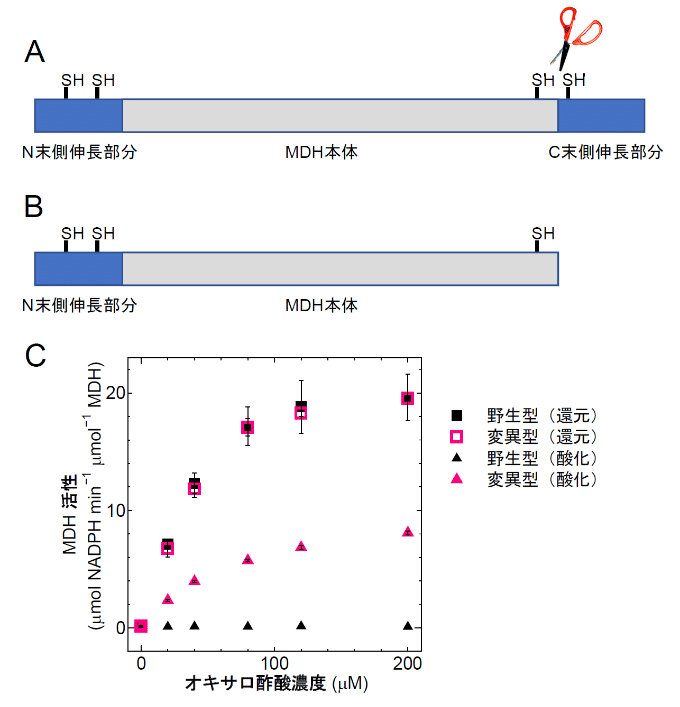
図2. MDH変異タンパク質と活性
そこで、ゲノム編集技術を用いて、上の組換え体タンパク質の実験と同じようにシロイヌナズナ葉緑体のMDHのC末端側のスイッチが働かないように遺伝子を書き換えた植物を作成した。こうして得られたMDHに変異を持った植物(MDH変異株)を育てて、緑葉から酵素を粗抽出してMDH活性を測定した。野生株の植物でも同様にMDHを粗抽出して比較したところ、レドックススイッチを壊したMDH変異株の酵素は、夜間でも高い活性を維持していることがわかった。
次に、野生株とMDH変異株の生育を光条件を変えて調べた。通常の長日(16時間明所/8時間暗所)の光条件では、生育にはほとんど違いがみられなかった。しかし、短日条件(8時間明所/16時間暗所)、超短日条件(4時間明所/20時間暗所)、光強度が強弱で大きく変動する変動光条件など植物の生育に不利な環境条件に置くと、変異株の生育の方が明らかに悪くなることがわかった(図3)。しかし、このような生育に影響する条件でも、クロロフィルの合成や実験室条件で測定した光合成活性には顕著な違いがみられなかったので、光条件による生育不全は、光合成反応よりも下流の細胞内の代謝機能の違い、すなわち還元力の蓄積と消費によって引き起こされていると予想した。

図3. 異なる光環境でのMDH変異株の生育
これを確認するために、埼玉大学の川合真紀教授のグループの協力を得てMDH変異株の代謝産物のメタボローム解析[用語10]をしたところ、リンゴ酸バルブ周辺の代謝産物量に顕著な変化が見られた。さらに、MDH変異株のNADとNADPの変動を調べてみると、細胞質側の還元力であるNADHが夜間には野生型よりも高くなっていることがわかった。このような還元力の分配バランスの変化が植物体の生育に直接影響するものと思われる。
今後の展開
本研究は、2020年のノーベル化学賞を受賞したCRISPR/Cas9を用いたゲノム編集技術を緑色植物の遺伝子の書き換えに用いることで、実現した。本研究により、緑色植物において、特に自然界のように光条件が常に変化する環境ではレドックス制御が重要であるということを明確に証明することができたことで、今後、植物のレドックス制御の研究はますます注目され、研究が加速するであろう。さらに、植物の代謝に直接かかわる制御機構を自由に操ることができれば、今後、農作物や光合成機能を利用したカーボンニュートラルに関わる様々な産業分野にも貢献できるものと期待している。
付記
本研究は、科学研究費助成事業 新学術領域研究「新光合成:光エネルギー変換システムの再最適化」(計画班代表:久堀徹教授)、東京工業大学情報生命博士教育院、および、World Research Hub Initiative(WRHI)の支援を受けて行われた。
- 用語説明
[用語1] レドックス制御機構 : 酸化還元状態に応じて、タンパク質分子の持っているジスルフィド結合(用語5)の形成解離を制御することにより、そのタンパク質の酵素活性を調節する分子機構。タンパク質の翻訳後修飾のひとつ。
[用語2] NADP-リンゴ酸脱水素酵素(MDH) : リンゴ酸脱水素酵素(MDH)は、リンゴ酸とオキサロ酢酸の相互変換を触媒する重要な酵素で、NADPを補酵素として用いるものをNADP-MDH、NADを補酵素として用いるものをNAD-MDHと呼ぶ。光合成生物では、還元力のやり取りに重要な働きをするほか、トウモロコシなどC4植物と呼ばれる特徴的な光合成を営む植物では、光合成反応そのものに重要な役割を持っている。また、NAD-MDHは、細胞呼吸においてクエン酸回路の重要な酵素のひとつである。
[用語3] NADPH : ニコチンアミドアデニンジヌクレオチドリン酸(NADP)の還元型。他の分子と電子のやり取りをすることで、酸化型のNADP+と還元型NADPHの間で相互変換する。光合成生物では、NADPHは炭素固定反応の還元力供給源となる。リン酸が結合していないものは、ニコチンアミドアデニンジヌクレオチド(NAD)という。
[用語4] 光合成電子伝達系 : 葉緑体チラコイド膜に局在し、水を分解し酸素発生と電子の生成を行う光化学系II、NADP+の還元を行う光化学系I、および、この両者をつなぐ電子伝達タンパク質であるシトクロムb6f複合体、という3つの主要なタンパク質複合体で構成されている。この3者の働きによって、水が分解されて酸素が発生し、還元物質であるNADPHを生成する。この反応の過程で、チラコイド膜のストロマ側からルーメン側(内腔側)にプロトンが輸送され、これによってチラコイド膜の内外に形成されるプロトンの電気化学ポテンシャル差(簡単に言えばプロトンの濃度勾配と膜電位の差の合計)を利用して、エネルギー物質であるATPが合成される。
[用語5] ジスルフィド結合 : システインの側鎖であるチオール基は、近接(通常4-5 Å以内)していて適当な酸化剤が作用すると共有結合を作る。これをジスルフィド結合と呼ぶ。
[用語6] プロテオミクス技術 : 生物の持っている遺伝子情報が解読されるようになったことで、細胞内に発現しているすべてのタンパク質の情報が一挙に入手できるようになった。この情報を用いて、細胞内など生体内にあるすべてのタンパク質の特徴などを網羅的に解析することをプロテオミクス(あるいはプロテオーム解析)と呼ぶ。
[用語7] N末端側とC末端側 : タンパク質は、20種類のアミノ酸がペプチド結合で連結したポリマーである。個々のアミノ酸は、(アミノ基)–(α-カーボン)–(カルボキシル基)で構成されていて、あるアミノ酸のカルボキシル基と次のアミノ酸のアミノ基の間の結合がペプチド結合である。したがって、アミノ酸ポリマーの両端は、アミノ基(NH2-)とカルボキシル基(-COOH)となるが、慣例上、アミノ基は左側に、カルボキシル基を右側に描く。そこで、タンパク質の方向をN末端側、C末端側と読んで区別する。
[用語8] Cys : システインというアミノ酸。α-カーボンには、アミノ基とカルボキシル基の他、水素原子1個と、側鎖が結合している。システインの側鎖は、SH-CH2-である。このSH部分は通常チオール基と呼ばれる。
[用語9] 組換え体タンパク質 : タンパク質をコードする遺伝子を発現ベクターと呼ばれるDNAに組み込み、このDNAをそのタンパク質を持っている生物とは別の生物の細胞内に導入してタンパク質を発現させたとき、得られるタンパク質を組換え体タンパク質(あるいは組換えタンパク質)という。発現させる細胞としては、大腸菌や酵母がよく用いられる。あらかじめ、タンパク質をコードする遺伝子を人為的に書き換えることで、アミノ酸配列に任意の変異を導入することができるので、この技術を用いると個々のアミノ酸の機能解析を行うことができる。
[用語10] メタボローム解析 : 細胞内に含まれる糖、アミノ酸、脂肪酸、有機酸などの代謝産物を質量分析によって網羅的に定量解析すること。
- 論文情報
| 掲載誌 : | Proceedings of National Academy of Science USA |
|---|---|
| 論文タイトル : | Redox regulation of NADP-malate dehydrogenase is vital for land plants under fluctuating light environment |
| 著者 : | Yuichi Yokochi, Keisuke Yoshida, Florian Hahn, Atsuko Miyagi, Ken-ichi Wakabayashi, Maki Kawai-Yamada, Andreas P. M. Weber, Toru Hisabori |
| DOI : | 10.1073/pnas.2016903118 |
- 光合成における新しい電子伝達タンパク質を発見|生命理工学系 News
- 細胞内の状態を可視化するセンサーの開発|生命理工学系 News
- 植物の酸化還元状態をリアルタイムで検知|生命理工学系News
- 高感度な酸素センサータンパク質を開発|生命理工学系News
- 植物はどのようにして眠るのか|生命理工学系News
- ラン藻による有用物質の大規模生産に道を拓く―高価な誘導剤使わずに遺伝子発現を誘導するネットワークを構築―|生命理工学系News
- 葉緑体機能の制御に重要な新たな還元力伝達経路―二つの経路の協調が光合成や生育に必須―|生命理工学系News
- 藻類の「眼」が正しく光を察知する機能を解明|生命理工学系News
- 藻類を使ったアンモニア生産の可能性―ラン藻の遺伝子発現を制御して放出させることに成功―|東工大ニュース
- 生体内のタンパク質の酸化還元状態を可視化 ―DNAを着脱自在にした修飾化合物を利用して総合的分析を実現―|東工大ニュース
- 久堀・若林研究室
- 研究者詳細情報(STAR Search) - 久堀徹 Toru Hisabori
- 科学技術創成研究院 化学生命科学研究所
- 科学技術創成研究院(IIR)
- 生命理工学院 生命理工学系
- Heinrich Heine University Düsseldorf
- 研究成果一覧
お問い合わせ先
東京工業大学 科学技術創成研究院 化学生命科学研究所
教授 久堀徹
E-mail : thisabor@res.titech.ac.jp
Tel : 045-924-5234 / Fax : 045-924-5268
おすすめ記事
-
2026.07.17
-

2026.07.13
-

2026.06.30
-
2026.06.23
-
2026.06.23
![]()