生命理工学系 News
光合成の安全装置「DLDG1」の役割を解明
植物が強い光から身を守る仕組みに新たな知見
ポイント
- 光ストレスから植物を守る仕組みを解析し、新たな視点・知見を提供
- DLDG1と呼ばれるタンパク質が葉緑体内pH調節と光防御に関与
- 強光に強い作物開発への応用に期待
概要
私たちの暮らしを支える植物は、光合成によって太陽の光エネルギーを使って有機物を作り出しています。しかし、光が強すぎると、逆に植物の体内では有害な活性酸素が発生し、細胞を傷つける恐れがあります。こうしたダメージを防ぐため、植物は「余分な光エネルギーを熱として逃がす」仕組みを備えており、これを熱放散[用語1]と呼びます。
本研究では、この熱放散の調節に関わるDLDG1[用語2]というタンパク質に注目しました。DLDG1は葉緑体の包膜にあるイオンの出入りを調節する装置(輸送体)で、これまでに、DLDG1が欠損すると熱放散が過剰に働くことが知られていました。しかし、DLDG1がどのように熱放散の仕組みとつながっているのか、はっきりと分かっていませんでした。
東京科学大学(Science Tokyo) 生命理工学院 生命理工学系の増田真二教授(生命理工学コース 主担当)らは今回、DLDG1を欠損させた植物と、葉緑体ATP合成酵素[用語3]の働きに異常があるhope2変異体[用語4]をかけ合わせ、dldg1-hope2 二重変異体を作製しました。この実験により、DLDG1が熱放散の強さや葉緑体内のpHバランスを調整する上で、ATP合成酵素との連携によって重要な役割を果たしていることが分かってきました。
本研究は、植物が環境ストレス(強光、乾燥、温度変化など)に適応するための「光の安全弁」の働きに、新たな視点を提供するものです。今後、作物の光合成効率やストレス耐性を高める研究への応用も期待されます。
研究成果は8月26日(現地時間)に「Plant Physiology」オンライン版に掲載されました。
背景
植物は、太陽光を利用して有機物を作り出しています(光合成)。しかし、強すぎる光が与えられると光合成で使われる電子が過剰になり、有害な活性酸素が生み出され細胞にダメージを与える可能性があります。このため植物は、過剰な光エネルギーを熱として放出する「熱放散」という防御機構を備えています。熱放散は、葉緑体内部のpH(酸性度)の変化により素早く起動しますが、このpHバランスを制御する仕組みには未解明の部分が多く残されています。中でも、「DLDG1」と呼ばれるタンパク質が熱放散に関与していることは知られていましたが、その具体的な働きはよく分かっていませんでした。
研究成果
今回、増田教授らの研究グループは、モデル植物シロイヌナズナを用いて、DLDG1の働きを詳しく調べました。DLDG1は葉緑体の包膜に存在するタンパク質で、水素イオンを出し入れすることで葉緑体内のpHを調節していると考えられています。これまでの研究で、DLDG1が欠損すると熱放散が過剰に働くことが分かっていましたが、その理由ははっきりしていませんでした。
本研究では、DLDG1を欠損させた変異体(dldg1)に、ATP合成酵素の調節に異常を持つhope2変異体をかけ合わせ、dldg1hope2二重変異体を作製し、光合成活性の比較を行いました。その結果、ストレス(変動光)下での光合成活性がhope2変異体において低下することが示され、dldg1hope2二重変異体ではより大きく低下することが分かりました(図1)。一方、hope2変異体では熱放散の働きが弱く、葉緑体内のpHバランスも乱れていましたが、dldg1の変異が加わることで、これらの異常が相殺され、熱放散が強化されることも分かりました。またdldg1の欠損により過剰となる熱放散が、hope2変異が加わることで相殺され、熱放散が弱まることも分かりました。つまり、DLDG1がATP合成酵素の働きと密接に関わりながら、葉緑体内のpHバランスを制御し、それによって熱放散の強さを調整している可能性が示されました。
この結果は、DLDG1が単独で働いているのではなく、葉緑体ATP合成酵素と連携することで、光の性質に応じた精密なpH調整を行っていることを示唆しています。DLDG1の働きを遺伝学的に抑制したことで、ATP合成酵素の調節異常を部分的に補えることが明らかになったのは、光合成調節メカニズムの理解を深める上で大きな進展です。
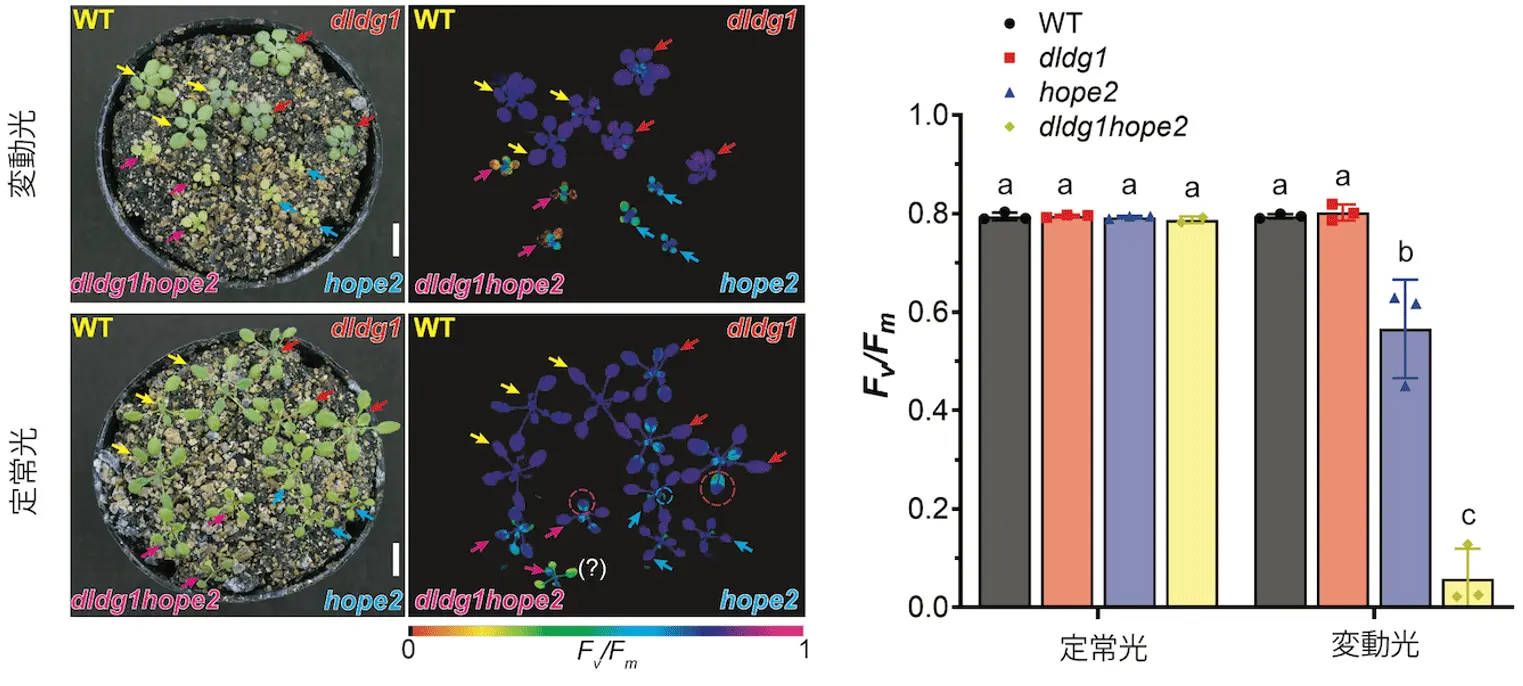
図1.(左)光の強さが数分ごとに変動する光(変動光)と全く変動しない光(定常光)で育てた野生型(WT)、dldg1変異体、dldg1hope2変異体、hope2変異体の写真。(右)光合成効率を反映するFv/Fm値。植物にとってストレスとなる変動光で育てると、hope2変異により低下したFv/Fm値が、dldg1の変異の追加によりさらに強く低下することが分かる。左図における矢印は、Fv/Fm値の定量化に用いた葉を示す。また、赤点線で囲んだ箇所はdldg1変異により薄緑色になりFv/Fm値が低くなった典型的な葉を示す。
社会的インパクト
植物の光合成効率とストレス耐性は、農業の生産性や作物の環境適応力に直結します。本研究で得られた知見は、強光や乾燥などの環境ストレスに強い作物の開発につながる可能性があり、将来の安定的な食糧生産や持続可能な農業への貢献が期待されます。
今後の展開
今後は、DLDG1がどのようにATP合成酵素と連動してpH調節を行うか、その分子メカニズムの解明を進めます。また、他の植物種における同様の仕組みの普遍性を調べることで、光合成制御の応用展開を視野に入れた研究を推進していきます。
- 付記
本研究は、コペンハーゲン大学 植物環境科学科のMichael Palmgren教授、神戸大学 大学院農学研究科の三宅親弘教授、理化学研究所 環境資源科学研究センターの佐藤諒一博士と共同で実施されました。またJSPS科学研究費助成事業(22K06276)の支援を受けて実施されました。
- 用語説明
[用語1] 熱放散:植物や藻類が持つ吸収した光エネルギーを熱として消費する機構。特定のカロテノイド分子が関わると考えられている。
[用語2] DLDG1:葉緑体の包膜に存在し、包膜を隔てた水素イオンの輸送を制御すると考えられているタンパク質。酸素発生型の光合成を行う植物や藻類のみに保存されている。Day Length-dependent Delayed-Greening1の略称で、各単語の頭文字をとってDLDG1と名付けられた。
[用語3] ATP合成酵素:膜を隔てたpH差(水素イオンの濃度差)を利用して、細胞のエネルギー通貨として働くATPを合成する酵素。
[用語4] hope2変異体:葉緑体で働くATP合成酵素の一部に特定のアミノ酸変異をもった植物体。この変異を持つATP合成酵素は、ATP合成の際に必要な膜内外pH差が、通常より大きいことが分かっている。
- 論文情報
| 掲載誌: | Plant Physiology |
|---|---|
| タイトル: | Chloroplast envelope-localized DLDG1 modulates H+ translocation across thylakoid membranes via plastidial ATP synthase |
| 著者: |
Mai Duy Luu Trinh1,2, Elham Esmailpourmoghadam3, Ryoichi Sato1,4, Chikahiro Miyake5, Michael Palmgren2, Shinji Masuda1,3 1東京工業大学 生命理工学院 2コペンハーゲン大学 植物環境科学科 3東京科学大学 生命理工学院 4理化学研究所 環境資源科学研究センター 5神戸大学 大学院農学研究科 |
| DOI: |
10.1093/plphys/kiaf373 |
研究者プロフィール
増田 真二 Shinji MASUDA
東京科学大学 生命理工学院 生命理工学系 教授
研究分野:植物生理学、光合成科学

関連リンク
- 太古の昔、生命を育んだ海は「緑色」だった!?|生命理工学系 News
- 生物は硫化水素を有効利用して生きている|生命理工学系 News
- 硫化水素が細菌の抗生物質耐性を高める仕組みを解明|生命理工学系 News
- 窒素不足の土壌でも“植物バイオマス”を増やせる葉緑体の働きを解明|生命理工学系 News
- 光合成における新しい電子伝達タンパク質を発見|生命理工学系 News
- 後生動物細胞からの内生グアノシン4リン酸(ppGpp)の検出に成功|生命理工学系 News
- 光合成の明反応と暗反応を協調させる仕組みを解明|生命理工学系 News
- 硫化水素に応答して遺伝子発現を調節するタンパク質を発見―硫化水素バイオセンサーの開発に道―|生命理工学系 News
- 葉緑体が植物の成長を制御する新たな仕組みを発見―細胞内共生した細菌の宿主細胞制御戦略―|旧・東京工業大学
- 高等植物の雄しべ発達過程を制御する植物ホルモン輸送体を発見|旧・東京工業大学
- 遺伝子発現を光で自在にコントロールする新技術を開発|旧・東京工業大学
お問い合わせ
東京科学大学 生命理工学院 生命理工学系
教授 増田 真二
Tel 045-924-5737
Fax 045-924-5823
Email shmasuda@bio.titech.ac.jp
おすすめ記事
-

2026.07.23
-

2026.07.22
-

2026.07.22
-
2026.07.17
-

2026.07.13
![]()