生命理工学系 News
シアワセモは強い光から逃げずに防御する
4細胞緑藻シアワセモの、近縁藻類とは異なる生存戦略を発見
要点
- 強い光は、光合成生物にとって生存に関わる脅威となる。緑藻類の多くは、強い光を感じると泳ぐ方向を変えて逃げる。今回、「4細胞で1個体」の世界最小細胞数の多細胞生物・緑藻テトラバエナ(和名:シアワセモ)は、この泳ぐ方向を変える機能が低いかわりに、強い光エネルギーを熱として捨てる光防御能力が高いことが分かった。
- 泳ぐのが下手なテトラバエナは、根を張って動けない陸上植物に似た生存戦略をとっていると考えられる。
概要
東京工業大学 生命理工学院 生命理工学系の丹野明日翔大学院生(研究当時)と同大 科学技術創成研究院 化学生命科学研究所の若林憲一准教授(ライフエンジニアリングコース主担当)、東京大学大学院 理学系研究科の野崎久義准教授(研究当時)、法政大学 自然科学センターの植木紀子教授らの研究グループは、細胞が4つしかない多細胞緑藻「テトラバエナ(和名:シアワセモ)」が、強い光刺激に対して、近縁の緑藻類とは異なる生存戦略をとっていることを明らかにした。
テトラバエナは繊毛[用語1]を使って水中を泳ぐ、緑藻綱ボルボックス目に属する藻類だ。緑藻などの光合成生物にとって光は重要なエネルギー源だが、強すぎる光は脅威となるため、生存に適した光環境を求めて遊泳する「光反応行動」は、遊泳性をもつ藻類にとって必須である。
研究グループはこれまでに、緑藻綱ボルボックス目に属する藻類であるクラミドモナスとボルボックスが、光刺激に対して、繊毛運動によって走光性[用語2]や光驚動反応[用語3]といった光反応行動を機敏に示すことを明らかにした。今回、テトラバエナにも同様の光刺激を与えたところ、驚いたことにテトラバエナは光受容機能が失われており、光反応行動を示さないことが分かった。その一方で、強すぎる光エネルギーを熱にして捨てる能力が非常に高いことを発見した。多細胞化進化の過程で光反応行動の能力を失いながらも、代わりに高い光防御能力を得たことで、自然界で生き残ってきたものと考えられる。
この成果は、PLOS ONEに10月26日にオンライン掲載された。
背景
テトラバエナ(和名:シアワセモ)は、淡水に棲む多細胞緑藻の一種である(図1左)。モデル単細胞緑藻クラミドモナス(和名:コナミドリムシ)によく似た細胞が4つ集まった体制をもつ(図1右)。4つの細胞が合わさっていること、またその形状が幸運の象徴である四つ葉のクローバーに似ていることなどから、「シアワセモ」という可愛らしい和名が付けられた。単細胞緑藻が複数寄り集まった姿ではないかとの議論があったが、2013年にれっきとした多細胞生物であることが証明された(Arakaki et al., 2013 PLOS ONE)。以来、「世界最小細胞数の多細胞生物」として知られている。
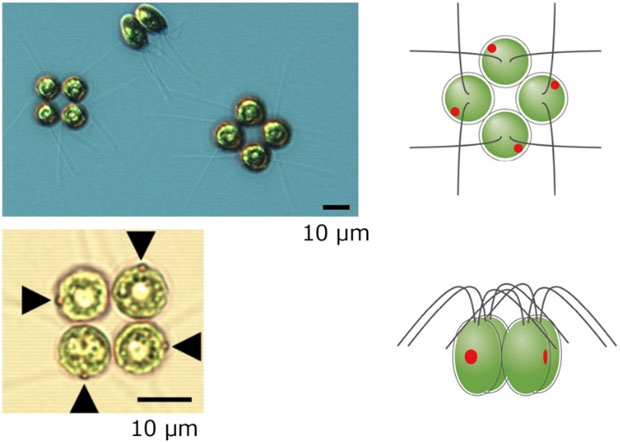
図1 テトラバエナの顕微鏡像(左)と模式図(右)。
DOI: 10.1371/journal.pone.0259138![]() より引用して改変。
より引用して改変。
分類上、テトラバエナは緑藻綱ボルボックス目に属する。この生物群は、クラミドモナス型の単細胞の祖先生物が約2億年前に多細胞化し、徐々に細胞数を増やしながら進化してきたと考えられている(図2)。単細胞のクラミドモナスから約1万細胞のボルボックスまでさまざまな細胞数の生物が現存しているため、生きている生物でありながら「段階的な多細胞化進化の歴史」の研究対象になるという、ユニークな生物群である。
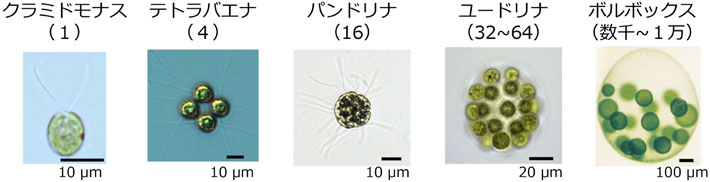
図2 さまざまなボルボックス目緑藻。カッコ内は細胞数。
ボルボックス目緑藻の細胞は、1つ1つはモデル単細胞緑藻クラミドモナスによく似ている。細胞に1つ「眼点」と呼ばれる赤い光受容装置をもち、2本の繊毛(鞭毛とも呼ばれる)を波のように動かして遊泳する。眼点は光を感じると外液からイオンを流入させることで細胞に「光を感じた」ことを知らせる(詳細は、以前の東工大ニュース「藻類の「眼」が正しく光を察知する機能を解明|生命理工学系 News」を参照)。光を感じた細胞の繊毛の動かし方が変わることで、緑藻は光反応行動を起こす。クラミドモナスは、2本の繊毛を打つ強さのバランス変化によって走光性を、波の形を変えることによって光驚動反応を示す。一方、ボルボックスは、繊毛を動かす面を回転させること、および球体のどの位置でその反応を起こすかを変化させることによって、走光性と光驚動反応を示す(図3)(詳細は、以前の東工大ニュース「ボルボックスの鞭毛が機能分化していることを発見|生命理工学系 News」を参照)。ボルボックスは、多細胞化の過程で、その巨体でも光反応行動を示すことができるように、クラミドモナス型祖先単細胞生物から繊毛の制御方法を変えたと考えられる。
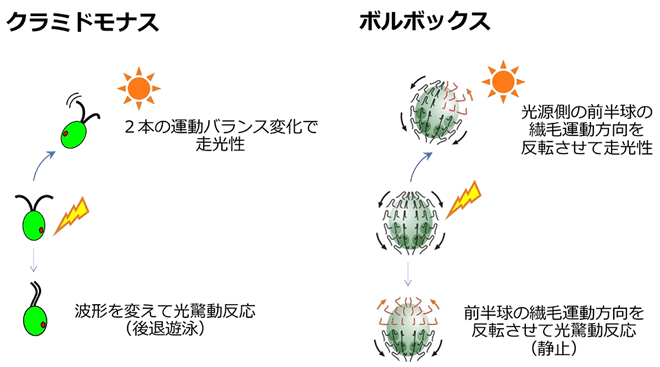
- 図3クラミドモナスとボルボックス繊毛制御方法の違い。クラミドモナスは通常は繊毛を平泳ぎのように動かして前進遊泳する。弱い光を感じると2本の繊毛の打つ強さのバランスを変えて遊泳方向を変え、走光性を示す。強い光を感じると繊毛の波の形を平泳ぎ型からドルフィンキック型に変えて光驚動反応(後退遊泳)を示す。ボルボックスは通常各細胞から生える2本の繊毛を両方とも後端方向に向かって打つことで前進遊泳する。光を感じると繊毛を打つ面を回転させるが、その角度は前端で約180度、赤道付近で約90度、後端でほぼ0度と前後軸に沿った勾配がある。つまり、前半球では運動方向をほぼ反転させる。弱い光を感じると光源側かつ前半球の繊毛が反転し、前後軸に対して光源側と反対側で推進力の不均衡が生じ、遊泳方向を変えて走光性を示す。強い光を感じると前半球の繊毛が反転し、前半球と後半球の生み出す力が釣り合って光驚動反応(静止)を示す。
研究成果
研究グループは、この多細胞化に伴う繊毛制御方法の変遷の初期過程を探るため、ボルボックス目の最初の多細胞種であるテトラバエナがどのように光反応行動を示すのかを調べた。すると、驚いたことに、光を当ててもテトラバエナは全く泳ぎ方を変えなかった。走光性も、光驚動反応も示さなかったのである(図4、動画)。その原因は、眼点にあった。テトラバエナ細胞にもクラミドモナスやボルボックスと同様の眼点は存在するが、光刺激を与えてもイオンの流入を全く起こさなかった。
しかし、テトラバエナは秒単位での走光性や光驚動反応は示さなかったものの、分単位でゆっくりと光に向かって集まる「光集合」と呼ばれる行動を示した。この行動は光合成に依存するが、光合成を出発点とするシグナル経路がどのようにしてゆっくりとした方向性のある運動を生み出すのか、そのメカニズムは謎であり、今後の課題である。

- 図4クラミドモナスとテトラバエナの走光性検定実験。それぞれの培養液をペトリディッシュにいれ、右側から眼点の光受容体がよく反応する緑色の光を照射した。クラミドモナスは正の走光性を示して右側に集まるが、テトラバエナは集まらなかった。
DOI: 10.1371/journal.pone.0259138 より引用して改変。
より引用して改変。
直後に繊毛の運動波形を変換して後退遊泳、
つまり光驚動反応を示した。動画は1/20倍速。
DOI: 10.1371/journal.pone.0259138![]() より引用
より引用
光合成生物にとって、光はエネルギー源であるが、強すぎる光は脅威になる。例えば、光合成反応が飽和しているときにさらに強い光を浴びると、光合成の反応場において活性酸素種が産生される。これによってタンパク質や脂質などが変性し、やがては細胞死に至る。光合成生物は、この光障害を避けるために多様な光防御システムを備えているが、その1つに、余分な光エネルギーを熱として捨てる熱放散機構[用語4]がある。陸上植物の葉緑体はいつでもこの機構を使うことができるが、クラミドモナスなどの緑藻の葉緑体は、この機構を誘導するタンパク質を発現させるという「下準備」ができて初めて作動する。クラミドモナスやボルボックスが示す光反応行動も、強すぎず弱すぎない適切な光環境に泳いで移動するための光防御システムの一環であると考えられている。
研究グループは、テトラバエナが光反応行動を示さないのは、他の光防御システムが発達しているからではないかと予測し、熱放散機構に着目したところ、テトラバエナの熱放散機構の能力はクラミドモナスよりも顕著に高く、かつ下準備を要せずいつでも発現可能であった(図5)。
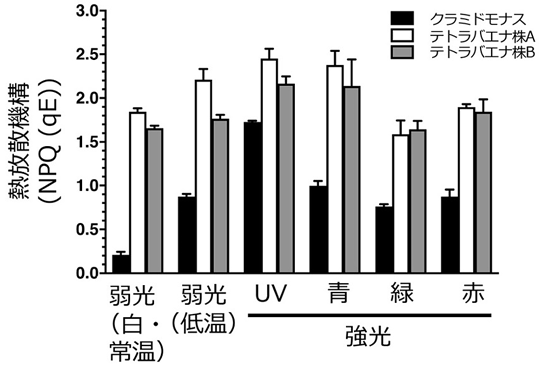
- 図5クラミドモナスとテトラバエナの熱放散機構。クラミドモナスは常温の弱光では値が低く、低温処理や強光照射などによって機構がONになり、値が高くなる。一方、テトラバエナは温度や光条件を問わずいつでも、かつクラミドモナスよりも顕著に高い値を示した。
DOI: 10.1371/journal.pone.0259138より引用して改変。
4細胞が横に並ぶ体制になったテトラバエナは、クラミドモナスやボルボックスのように上手に遊泳方向の舵取りをすることができない。そのため、光反応行動を半ば捨て、その代わり、根を下ろしたらその場から動けない陸上植物によく似た「常に熱放散機構ON」という生存戦略をとったと予想される(図6)。このことは、テトラバエナが生息する場所が浅い池や極地の水たまりなど、日光から逃げにくい場所であることとも関係すると考えられる。
光反応行動の生理的意義は、これまではっきりとした証明実験がなかった。今回、迅速な光反応行動を示せないテトラバエナの熱放散能力が高かったことは、光反応行動が光防御システムの一環であるとの考えを強めるものと推測できる。
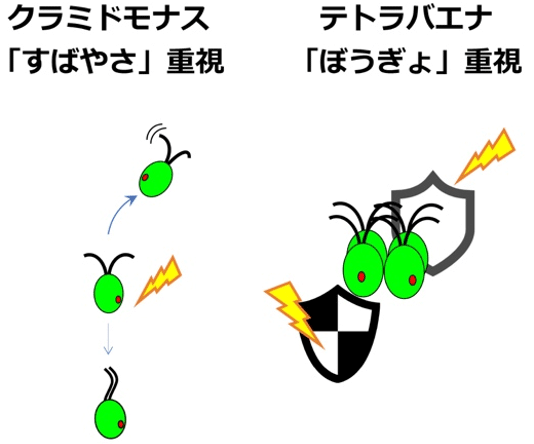
- 図6クラミドモナスとテトラバエナの生存戦略の違い。クラミドモナスは強光を浴びると負の走光性や光驚動反応によって光から逃げ、その間に熱放散機構をONにする。一方、テトラバエナは強光から逃げられない代わりに、常に熱放散機構をONにして備えている。
今後の展開
他の遊泳性藻類で今回と同様の実験を行うことによって、今回テトラバエナとクラミドモナスの比較で判明した光反応行動能力と熱放散能力の負の相関関係が、他の生物にも適用できるのかどうかを明らかにする。これにより、地球上の重要な一次生産者である藻類の生存戦略の一端の解明につなげる。また、テトラバエナの「常時熱放散機構ON」のしくみを明らかにすることによって、バイオ燃料等を産生する有用藻類を屋外で大量培養する際の効率化に貢献できるものと期待される。
- 付記
本研究は科学研究費補助金(15H05599, 16H06556, 17K07370, 19H03242, 20H03282, 20K21420, 21H00420, 21K06295)、大隅基礎科学創成財団、ダイナミック・アライアンスの援助を受けて行った。
- 用語説明
[用語1] 繊毛 : 真核生物細胞から生える毛状の運動する細胞小器官。本数が少なく長いものを鞭毛、本数が多く短いものを繊毛と呼ぶが、本質的には同じものであり、緑藻のものは繊毛と用語統一されつつある。精子のように細胞の推進力を生み出したり、気管上皮のように細胞の周囲に水流をつくったり、生体にとって重要な機能をもつ。ヒト体内には脳室、気管、卵管、精子などに運動性鞭毛・繊毛が存在する。それらの運動異常によって生じる疾患は原発性不動繊毛症候群と呼ばれる。
[用語2] 走光性 : 生物が照射される光に反応して移動する性質。光源方向に近づく場合は正の走光性、離れる場合は負の走光性と呼ぶ。光走性(ひかりそうせい)と呼ばれることもある。
[用語3] 光驚動反応 : 生物が強い光強度変化に応答して、運動を止めたり、運動方向を逆転させたりする反応。光忌避反応と呼ばれることもある。
[用語4] 熱放散機構 : 光合成の反応場である光化学系複合体が、過剰な光エネルギーを浴びたときに複合体を再編し、光エネルギーを熱として発散させる機構。クロロフィル蛍光の非光化学的消光(Non-photochemical quenching)のqEクエンチングとして測定ができる。
- 論文情報
| 掲載誌 : | PLOS ONE |
|---|---|
| 論文タイトル : | The four-celled Volvocales green alga Tetrabaena socialis exhibits weak photobehavior and high-photoprotection ability |
| 著者 : | Asuka Tanno, Ryutaro Tokutsu, Yoko Arakaki, Noriko Ueki, Jun Minagawa, Kenjiro Yoshimura, Toru Hisabori, Hisayoshi Nozaki, and Ken-ichi Wakabayashi |
| DOI : |
- 光で繊毛運動を調節する新規タンパク質の発見|生命理工学系 News
- 細胞の運動を「10秒見るだけ」で細胞質ATP濃度がわかる|生命理工学系 News
- ボルボックスの鞭毛が機能分化していることを発見|生命理工学系 News
- 藻類の「眼」が正しく光を察知する機能を解明|生命理工学系 News
- 鞭毛モーターの規則的配列機構を解 -鞭毛を動かす " エンジン" が正しい間隔で並ぶ仕組み発見-|東工大ニュース
- 久堀・若林研究室
- 若林憲一 Ken-ichi Wakabayashi|研究者検索システム 東京工業大学STARサーチ
- 科学技術創成研究院 化学生命科学研究所
- 科学技術創成研究院(IIR)
- 生命理工学院 生命理工学系
- 東京大学大学院 理学系研究科
- 法政大学
- 研究成果一覧
お問い合わせ先
東京工業大学 科学技術創成研究院 化学生命科学研究所
准教授 若林憲一
E-mail : wakaba@res.titech.ac.jp
Tel : 045-924-5235 / Fax : 045-924-5268
おすすめ記事
-
2026.07.17
-

2026.07.13
-

2026.06.30
-
2026.06.23
-
2026.06.23
![]()