生命理工学系 News
藻類のオイル生産を制御する因子を同定
有用脂質生産の自在制御に向け大きな一歩
要点
- 藻類はリンや窒素などの欠乏時に細胞内にオイルを高蓄積
- 藻類で栄養欠乏時に起こるオイルの高蓄積を制御する制御因子を発見
- 制御因子の改変により、オイル生産を自在に制御する仕組みへの活用に期待
概要
東京工業大学 生命理工学院のNur Akmalia Hidayati(ヌル・アクマリア・ヒダヤティ)博士後期課程3年、堀孝一助教、太田啓之教授、下嶋美恵准教授、岩井雅子特任助教(共に生命理工学コース主担当)と京都大学 福澤秀哉教授、東北大学 大学院情報科学研究科 大林武准教授、かずさDNA研究所 櫻井望チーム長(現所属・国立遺伝学研究所)らの研究グループは、バイオ燃料をはじめとする有用脂質生産に活用が期待される藻類の一種「クラミドモナス[用語1]」で、リンや窒素の栄養欠乏時に起こるオイルの蓄積を制御する因子の同定に成功した。またこの制御因子は、特に栄養欠乏時の細胞内にオイルが大量に蓄積する時期に機能する主要な制御因子であることも突き止めた。
今回、種々の藻類で広く見られる栄養欠乏時のオイルの大量蓄積を制御する制御因子を見出したことで、明らかになった脂質蓄積の制御の機構や制御因子自体を、藻類で生産する有用脂質の種類や生産の時期を自在にコントロールするための仕組みづくりに活用することが期待される。
藻類はリンや窒素などの栄養欠乏時に細胞内にオイルを多量に蓄積することが広く知られている。この仕組みの解明が藻類で様々な有用脂質を自在に生産するための大きな手掛かりになると考えられていた。
研究成果は7月27日発行の英国科学雑誌「プラント ジャーナル(The Plant Journal)」に掲載された。
(注)この研究は、科学研究費基盤研究A、科学技術振興機構 産学共創プラットフォーム共同研究推進プログラム(OPERA) 「ゲノム編集による革新的な有用細胞・生物作成技術の創出」研究領域(研究総括:山本卓(広島大学教授))における研究の一環として行った。
研究の背景と経緯
これまで石油資源から生産されている様々な有用脂質を、光合成を行う藻類や植物で生産するバイオ燃料などで代替して製造し、石油資源への高い依存から脱却することが期待され、世界中で研究が進められている。中でも藻類は単位面積あたりの生産性が高いことや食用作物と競合しないという利点を持つ。
藻類が作り出すオイル(油脂、トリアシルグリセロール)は液体燃料として直接転用可能な原料となり、単位容積あたりのエネルギー効率も高いことから、ディーゼル燃料や航空燃料の代替として最適なバイオマスと期待されている。とりわけ、藻類はリンや窒素の欠乏時に細胞内に多量のオイルを蓄積することが広く知られており、その仕組みの解明に向けた研究が行われてきた。
藻類の栄養欠乏時におけるオイル蓄積を制御する仕組みを解明することができれば、その知見や制御を担う因子を活用することで、藻類で様々な有用脂質を適当な時期に自在に生産するシステムを構築することができると期待されている。しかし、特に栄養欠乏時のオイルの蓄積が顕著になる時期に油脂合成を制御する制御因子はこれまで全く同定されておらず、藻類の応用を見据えた基礎研究の推進が急務となっていた。
研究成果
太田教授らの研究グループは、モデル藻類のクラミドモナスを用いてオイル合成の最終段階を担う酵素「ジアシルグリセロールアシルトランスフェラーゼ(DGAT)[用語2]」の遺伝子の一つであるDGTT1[用語3]と同調的に遺伝子の発現が起こる転写因子の候補を網羅的に探索[参考1]した。その中から、特にリン欠乏条件移行後のオイルが大量に貯まる時期に強く発現する転写因子LRL1(Lipid Remodeling reguLator1)[用語4]の遺伝子を候補として見出し、解析をした。
LRL1の機能を明らかにするため、京都大学福澤研究室と共同で、遺伝子の発現が抑制された変異体を1ライン単離し、またクラミドモナスの遺伝子変異体のライブラリからLRL1の機能が抑制された変異体をもう1ライン単離して、それらの性質を詳細に解析した。その結果、変異体ではいずれもリン欠乏時にみられるオイルの蓄積が大きく抑制されていることが分かった。
さらにこれらの変異体では、栄養が十分ある際には細胞分裂が促進されて細胞のサイズが小さくなること、栄養が欠乏すると逆に細胞の増殖が抑制され、野生型に比べて細胞の色がやや薄緑色になることなどが分かった。またリン欠乏時に細胞内で不足したリンを生体膜のリン脂質から切り出して補い、リン脂質の代わりにリンを含まない糖脂質などを合成するリン欠乏時の膜脂質転換[用語5]と呼ばれる仕組みにも異常が起こっていることが分かった。
LRL1がこれらの現象に関わる遺伝子を直接制御しているかどうかを明らかにするため、タバコの葉を利用した遺伝子の一過的な発現系を用いて、藻類の転写因子が、藻類の膜脂質転換に関わる遺伝子を直接制御しているかどうかを解析した。その結果、LRL1は、クラミドモナスに存在する別の転写因子(bHLH2)と二つの転写因子の結合を促進するタンパク質(TTG1)と共同することで、リン欠乏時に起こるダイナミックな脂質代謝の変動を直接制御していることが明らかになった。
また、LRL1は、リン欠乏応答の早期に関わる転写因子として広く知られるPSR1[用語6]と同様な遺伝子を制御するが、PSR1とは異なり、リン欠乏のみならず窒素欠乏時にも発現が誘導されることから、栄養欠乏時の比較的後期に顕著にみられるオイルの蓄積や細胞分裂の抑制を制御する重要な制御因子であると考えられる。
今後の展開
今回の研究成果により、藻類の栄養欠乏時に広く見られるオイルの蓄積や膜脂質転換など脂質代謝の大きな変動(脂質転換)を制御する重要な制御因子の存在が明らかになった。今後、今回明らかになったLRL1の機能の仕組みやLRL1自体の活用により、藻類で大量生産が望まれる様々な有用脂質を必要な時期に自在に誘導蓄積する仕組みの構築が可能になることが期待される。
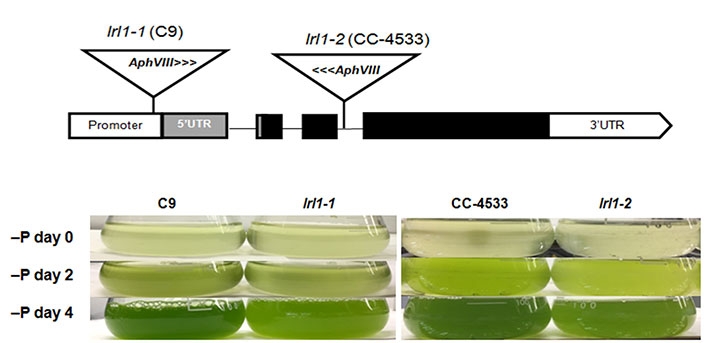
図1. LRL1変異体ではリン欠乏時の生育が抑制される。
上図:LRL1の二つの変異体、lrl1-1、lrl1-2のタグ挿入部位、下図:リン欠乏時の培養の様子。lrl1-1、lrl1-2では、いずれもリン欠乏時の細胞の増殖が抑制され、培養液がやや薄緑色になる。
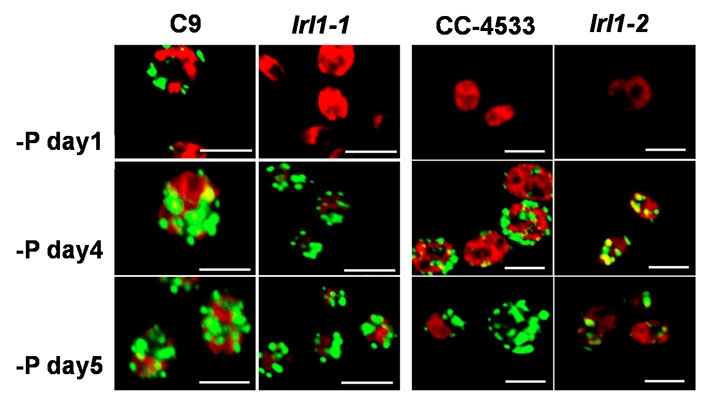
図2. LRL1の二つの変異体のオイルの蓄積と細胞サイズの変化
赤:葉緑体のクロロフィル蛍光 緑:油滴
リン欠乏条件(-P)への移行後、野生型(C9とCC4533)では細胞のサイズが肥大し、オイルの蓄積に伴い油滴の肥大や数の増加が起こるが、二つの変異体lrl1-1、lrl1-2では、オイルの蓄積が抑制され、細胞のサイズも小さい。
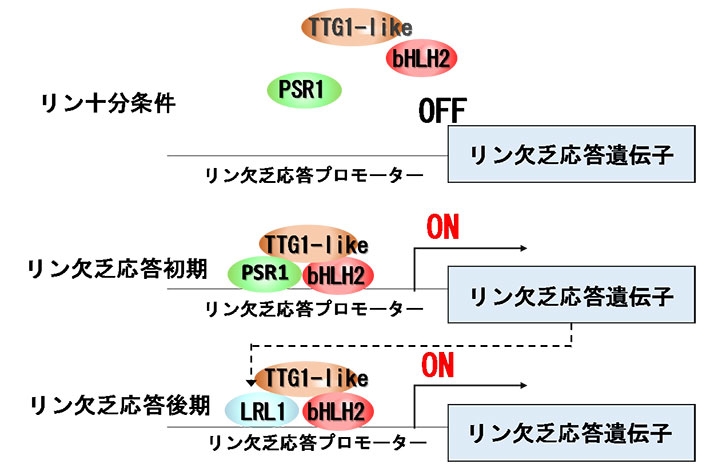
図3. リン欠乏時におけるLRL1の機能のモデル
LRL1は、他の転写因子bHLH2や、制御因子の連結に関わる因子TTG1と共同してリン欠乏応答遺伝子の発現誘導に直接働いていることが明らかになった。
- 用語説明
[用語1] クラミドモナス : モデル藻類として様々な研究に用いられる単細胞性の緑藻。窒素やリンの欠乏時に細胞内にデンプンとオイルを多量に蓄積することから、栄養欠乏時のオイル蓄積の研究のモデルとしても広く用いられている。
[用語2] ジアシルグリセロールアシルトランスフェラーゼ(DGAT) : 植物や藻類、動物など様々な生物でオイル(triacylglycerol,TAG)の合成の最終段階を担う重要な酵素。TAGの合成の前駆体であるジアシルグリセロール(グリセロール骨格に脂肪酸が2つエステル結合したもの)にさらにもう一つ脂肪酸を結合させる反応を触媒する。大きく分けてDGAT1とDGAT2の2つのタイプがある。
[用語3] DGTT1 : クラミドモナスには、5種類のDGAT2遺伝子が存在し、その中で、特に窒素やリンの欠乏時に遺伝子の発現が著しく上昇する遺伝子。
[用語4] LRL1(Lipid Remodeling reguLator1) : クラミドモナスで今回見出されたMYB型と呼ばれる転写因子の一種。MYB型の転写因子は藻類や植物、動物に広く存在することが知られており、それぞれの生き物で特定の代謝系の制御などを行うMYB型転写因子が多数存在する。それらの生き物では、MYB型転写因子をはじめとする種々の転写因子などの制御のネットワークにより、複数の代謝系や様々な生理現象が巧妙に維持されている。
[用語5] リン欠乏時の膜脂質転換 : 藻類や植物のリン欠乏時には細胞内に不足したリンを補うため、生体膜の主要構成成分であるリン脂質を分解して細胞内にリンを供給するとともに、リン脂質の代わりとしてリンを含まない糖脂質やベタイン脂質などを用いることで、リンの欠乏時への適応を行っている。また、リン欠乏時や窒素欠乏時には、細胞の増殖を抑制するとともに、栄養が十分に得られるようになった時に迅速に対応できるよう光合成で得られた余剰の炭素を貯蔵脂質(トリアシルグリセロール、オイル)として蓄積する。このような脂質代謝の変動は、生体膜とは異なり細胞質の脂質で起こるため、栄養欠乏時における膜脂質転換と合わせて「栄養欠乏時の脂質転換」と呼ばれている。
[用語6] PSR1 : リン欠乏時に起こる様々な現象を制御する中心的な制御因子。LRL1と同様MYB型の転写因子に属するが、LRL1とはサブグループが異なる。PSR1はR1型MYB、LRL1はR2R3型MYBのサブグループにそれぞれ属する。
- 参考情報
[1] 2016年に大林准教授、太田教授らが共同で開発した共発現データベースALCOdbを活用したものである。
Aoki Y, Okamura Y, Ohta H, Kinoshita K, Obayashi T. ALCOdb: Gene Coexpression Database for Microalgae. Plant Cell Physiology, 57, e3 (2016)
- 論文情報
| 掲載誌 : | The Plant Journal |
|---|---|
| 論文タイトル : | LIPID REMODELING REGULATOR 1 (LRL1) is differently involved in the phosphorus-depletion response from PSR1 in Chlamydomonas reinhardtii |
| 著者 : | Hidayati, Nur Akmalia; Yamada-Oshima, Yui; Iwai, Masako; Yamano, Takashi; Kajikawa, Masataka; Sakurai, Nozomu; Suda, Kunihiro; Sesoko, Kanami; Hori, Koichi; Obayashi, Takeshi; Shimojima, Mie; Fukuzawa, Hideya; Ohta, Hiroyuki |
| DOI : | 10.1111/tpj.14473 |
- 花を作る遺伝子の起源推定に成功 | 生命理工学系 News
- 大量のオイルを生産する“最強藻類”の秘密を解明 | 生命理工学系 News
- 葉緑体が植物の成長を制御する新たな仕組みを発見―細胞内共生した細菌の宿主細胞制御戦略― | 東工大ニュース
- 微細藻類にオイルをつくらせるスイッチタンパク質を発見―バイオ燃料生産実現に向けた基盤技術として期待― | 東工大ニュース
- 油脂高生産藻の脂質量と組成を改変する技術を開発―藻による油脂やバイオ燃料の生産性向上に期待― | 東工大ニュース
- 貧栄養土壌でも葉と根に油脂蓄積する植物を開発 | 東工大ニュース
- 高等植物の雄しべ発達過程を制御する植物ホルモン輸送体を発見 | 東工大ニュース
- 光合成反応の場を作る膜脂質の機能を解明 ―光合成膜脂質のこれまでの常識を覆す― | 東工大ニュース
- 藻類の栄養欠乏応答性プロモーターによる脂質蓄積強化を実現 | 東工大ニュース
- 藻類から陸上植物への進化をつなぐ車軸藻植物のゲノム配列を解読 | 東工大ニュース
- 太田啓之教授が2018年テリー・ガリアード・メダルを受賞 | 生命理工学系 News
- 太田・下嶋研究室 ―研究室紹介 #12― | 生命理工学系 News
- 太田・下嶋研究室
- 研究者詳細情報(STAR Search) - 太田啓之 Hiroyuki Ohta
- 研究者詳細情報(STAR Search) - 堀孝一 Koichi Hori
- 研究者詳細情報(STAR Search) - 下嶋美恵 Mie Shimojima
- 研究者詳細情報(STAR Search) - 岩井雅子 Masako Iwai
- 生命理工学院 生命理工学系
- 京都大学 大学院生命科学研究科 微生物細胞機構学分野
- 東北大学 大学院情報科学研究科
- 国立遺伝学研究所 生命情報・DDBJセンター
- かずさDNA研究所
- 研究成果一覧
お問い合わせ先
東京工業大学 生命理工学院 生命理工学系
教授 太田啓之
E-mail : hohta@bio.titech.ac.jp
Tel : 045-924-5736 / Fax : 045-924-5527
京都大学 大学院生命科学研究科 微生物細胞機構学分野
教授 福澤秀哉
E-mail : fukuzawa@lif.kyoto-u.ac.jp
Tel : 075-753-4298 / Fax : 075-753-9228
東北大学 大学院情報科学研究科
准教授 大林武
E-mail : obayashi@ecei.tohoku.ac.jp
Tel : 022-795-7161 / Fax : 022-795-7179
国立遺伝学研究所 生命情報・DDBJセンター
特任准教授 櫻井望
E-mail : sakurai@nig.ca.jp
Tel : 055-981-6895 / Fax : 055-981-9448
おすすめ記事
-

2026.07.23
-

2026.07.22
-

2026.07.22
-
2026.07.17
-

2026.07.13
![]()