生命理工学系 News
体内栄養状態を感知するmTORC1経路の活性制御機構を解明
リソソーム膜上におけるTSC2の選択的脱リン酸化がmTORC1の活性を厳密に制御する
ポイント
- 質量分析を活用して、mTORC1経路(図1)の構成タンパク質の細胞内濃度測定(iMPAQT)、リソソーム膜上タンパク質の網羅的同定(Lyso-BioID)を実施しました。
- 上記実測データを基に、mTORC1経路の数理モデルを構築して、mTORC1経路の活性制御機構を解明しました。
- インスリン刺激によって活性化されたAKTは、TSC2をリン酸化して、TSC1/2のブレーキ機能(GAP活性)を阻害するが、アミノ酸除去時では、TSC2のリン酸化レベルは、アミノ酸添加時と比べて有意に低いことを明らかにしました(図2、図5)。
- アミノ酸除去時にはTSC2はリソソーム膜上に移動してPP2Aによって選択的に脱リン酸化されることを明らかにした(図3、図4、図5D)。
- PP2A-TSC2を介したmTORC1経路の制御は、mTORC1がアミノ酸と増殖因子(インスリン)を同時に感知した場合のみ活性化することを保証するシステムであり、その破綻は、がんや、結節性硬化症などの疾患発症を惹起することも明らかにしました(図4、図5)。
概要
東京科学大学(Science Tokyo) 生命理工学院 生命理工学系(神奈川県立がんセンター兼任)の越川直彦教授(人間医療科学技術コース 主担当)は、愛媛大学 先端研究院 プロテオサイエンスセンター病理学部門の中村貴紀助教、増本純也教授、澤崎達也教授の研究グループ、東京大学 医科学研究所 武川睦寛教授、大阪大学 先端モダリティ・DDS研究センター 岡田雅人特任教授、同数理・データ科学教育研究センター 鈴木貴特任教授(常勤)、新潟大学 大学院医歯学総合研究科 松本雅記教授らとの共同研究で、栄養シグナル伝達の中心的役割を担うタンパク質複合体mTORC1の活性制御機構を解明することに成功しました。
mTORC1は、アミノ酸経路及びインスリン経路(AKT-TSC1/2-Rheb)によってリソソーム膜上で活性化し、体内の栄養状態に応じて、生体成分であるタンパク質、脂質、核酸などを合成するか、または分解するかを決定し、生命の恒常性維持を担います。mTORC1の機能破綻はがんや糖尿病など代謝性疾患を惹起するため、mTORC1の活性化は厳密に制御されますが、その詳細な分子機構は不明な点が多く残されています。今回我々は、近接ビオチン標識(Lyso BioID)などの実験手法と理論手法(数理解析)を駆使して、mTORC1シグナル経路の統合的理解を目指しました。その結果、インスリン刺激に伴うAKTキナーゼは、細胞内アミノ酸濃度に関係なく活性化されましたが、低アミノ酸レベルでは、AKTの基質分子でmTORC1経路のブレーキ役となるTSC2が、リソソーム膜上で脱リン酸化酵素PP2Aによって選択的に脱リン酸化されることを新たに見出しました。この脱リン酸化反応は、mTORC1がアミノ酸及びインスリンを同時に感知した場合のみ活性化することを保証する分子機構の1つであることを明らかにしました。さらに、結節性硬化症(指定難病158)及びがん症例の中に、TSC2のPP2A結合能を消失する遺伝子変異を見出しており、PP2A-TSC2によるmTORC1経路のブレーキ機能が破綻すると、上記のTSC2関連疾患の発症を惹起することも解明しました。
本研究成果は、9月2日10時(米国東部時間)に、米国の国際科学雑誌「Life Science Alliance」に掲載されました。
背景
ヒトの身体を構成する細胞は、増殖因子(例:インスリン)による刺激、細胞外からのアミノ酸などの栄養源あるいは、細胞内エネルギーを用いて、生体を構成するタンパク質、核酸、脂質などの高分子合成(同化作用)と、飢餓状態時には同高分子の分解(異化作用)とを、適切に制御することにより生命活動を維持しています。細胞が、栄養状態に応じて同化あるいは、異化反応いずれを誘導するか運命決定を担う鍵分子として、mTORC1(mTORタンパク質複合体1[用語1])が知られています。mTORC1は、種を越えて高度に保存されるリン酸化酵素mTORを核とするタンパク質複合体で、タンパク質、脂質、核酸などの生合成に関わる制御タンパク質をリン酸化することによって同化反応を促進する一方で、オートファジーやリソソーム合成に関わる分子をリン酸化することにより、生体高分子を分解して栄養源を産生する異化反応を抑制します(図1)。
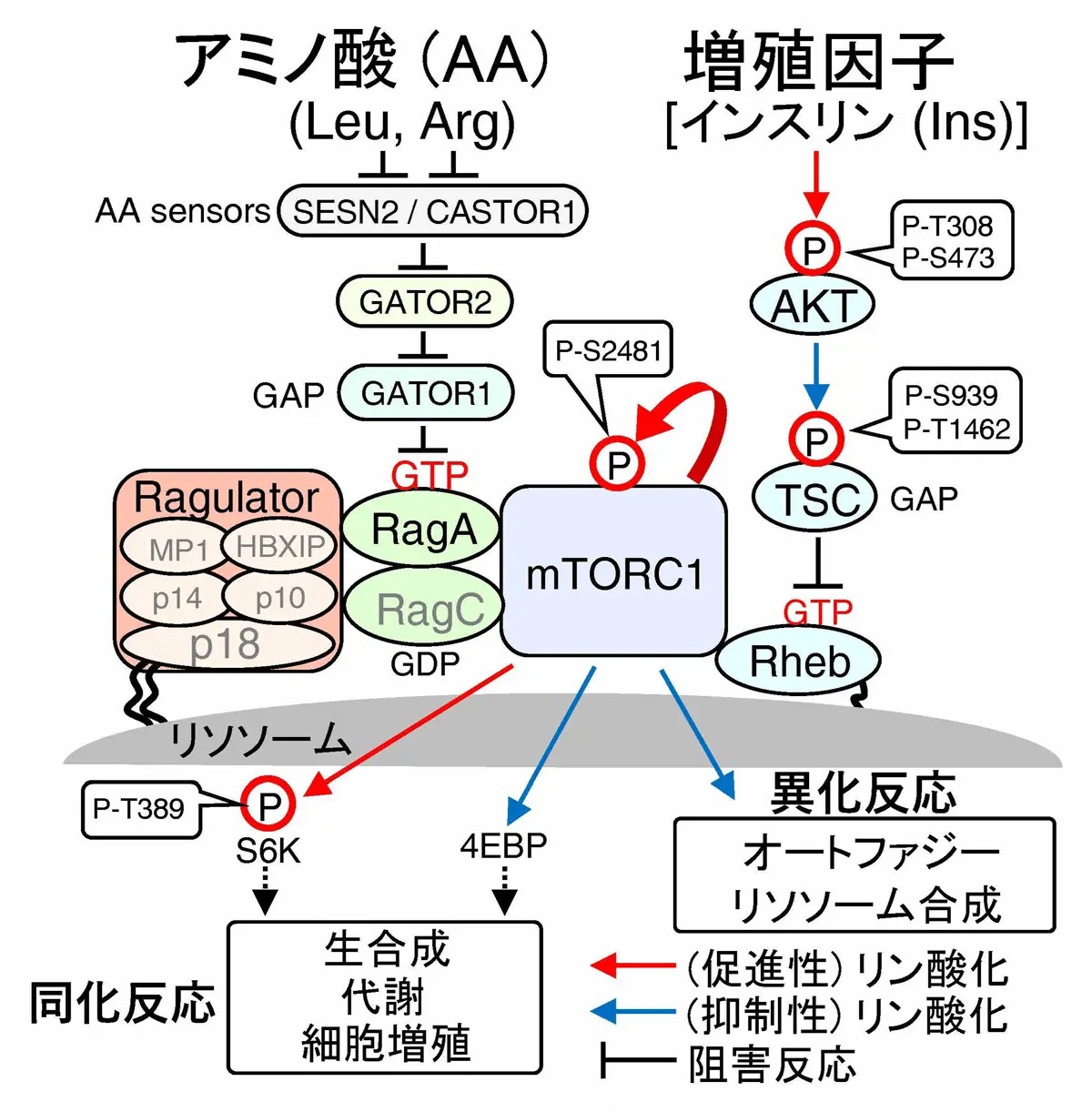
図1.mTORC1栄養シグナル経路
mTORC1はリソソーム[用語2]膜表面に分布する(活性型)RagGTPや(活性型)RhebGTPによって活性化されます。Rag及びRhebは、栄養シグナルが伝播されない場合には不活性化状態にあります。このため、mTORC1が活性化するには、まず、Rag及びRhebがアミノ酸感知経路(Ragulator-Rag)及び増殖因子(インスリン)感知経路(AKT-TSC1/2-Rheb)により活性化される必要があります。 Rhebの活性(RhebGTP)は、GAP活性を持つTSC1/2[用語3]によって不活性型(RhebGDP)に保たれていますが、TSC2がインスリン刺激によって活性化したAKTキナーゼにより、リン酸化されるとTSC2のGAP活性が抑制されるため、リソソーム膜上のRhebが活性化されます。 また、細胞が栄養素(アミノ酸)を感知すると、Ragulator (p18-p14-MP1-p10-HBXIP複合体)を介して、活性型RagGTPが、リソソーム膜上に集合します。Ragulator-RagGTPは、mTORC1のリソソーム移行を促進するため、リソソーム膜上でmTORC1が、活性化することも報告されています。
このようにmTORC1は、細胞内のアミノ酸濃度、あるいは、インスリン濃度を適切に検知して同化/異化反応のOn/Offを制御しているため、mTORC1栄養シグナル経路の調節システムの破綻は、がんや、糖尿病などの疾患発症を惹起することが知られています。例えば、インスリン感知(AKT-TSC1/2-Rheb-mTORC1)経路には、がんにおいて高頻度に変異が生じるがん遺伝子(AKT、mTOR)や、がん抑制遺伝子(PTEN、TSC1/2)が位置していることから、がんの発症や進展に関与する重要なシグナル伝達経路として、薬剤標的となっています。 mTORC1の活性制御機構については、近年世界中で盛んに研究が行われており、その全貌が明らかになりつつありますが、「アミノ酸経路、インスリン経路など複数の情報伝達を混線なく適切に制御するか」など、詳細な分子制御機構は、現在でも不明な点が多く残されています。
研究成果
研究グループは、細胞内タンパク質濃度の測定(iMPAQT[用語4])、リソソーム膜表面タンパク質の同定(Lyso-BioID[用語5])などの実験手法と、理論手法(数理解析)を駆使して、mTORC1栄養シグナル経路を、包括的に理解することを目指しました。その結果、インスリン感知(AKT-TSC1/2-Rheb)経路のAKTキナーゼは、細胞内アミノ酸濃度に関係なくインスリン刺激によって活性化されました(図2A)。一方AKTによるTSC2の(抑制性)リン酸化はアミノ酸添加条件に比べて、アミノ酸除去条件で顕著に低いことが分かりました(図2A)。さらに、TSC2リン酸化レベルが低い理由として、アミノ酸除去時には、TSC2がリソソーム膜上に移行して、脱リン酸化酵素PP2Aによって、選択的に脱リン酸化されることを新たに見出しました(図2B、図3、図4A、B、図5)。このリソソーム膜上におけるPP2AによるTSC2の脱リン酸化システム は、細胞がアミノ酸及びインスリンを同時に感知した場合にのみmTORC1が活性化することを保証する分子制御機構の1つであることを明らかにしました。さらに、結節性硬化症(指定難病158) 及びがん症例において、TSC2分子のPP2A-B56結合領域([L/M/F/I]-x-x-[I/L/V]-x-E-x)に、L366P(結節性硬化症)、E366K(大腸がん、卵巣がん)、E1583K(皮膚がん)などの遺伝子変異が生じていることを新たに見出しました(図4C、D)。この結果から、PP2A-TSC2によるmTORC1経路のブレーキ機能が破綻すると、上記のTSC2関連疾患の発症を惹起することも解明しました。
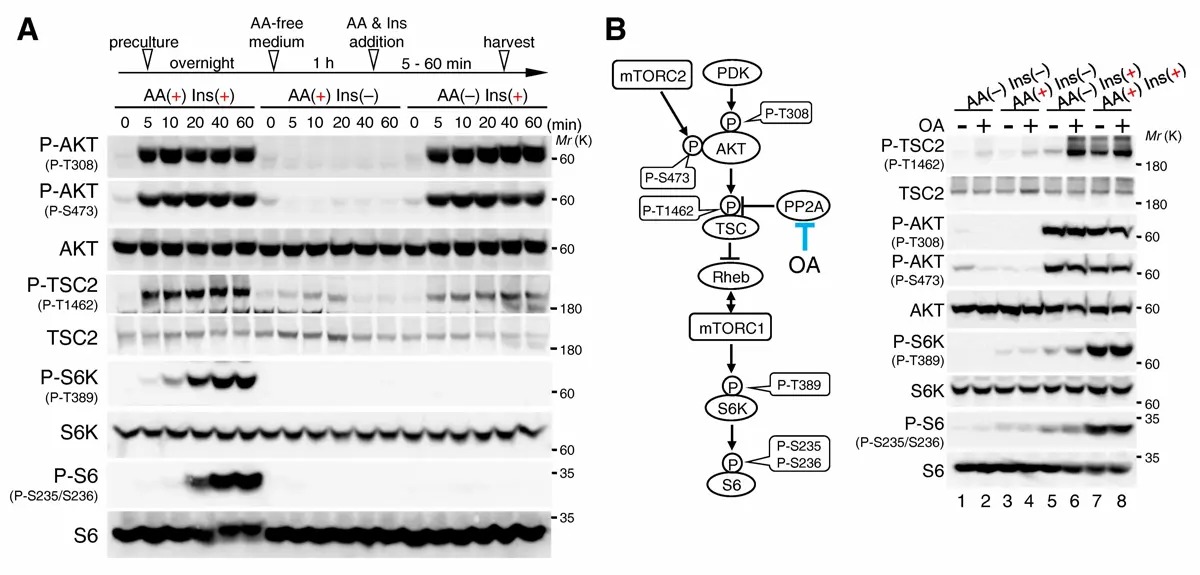
図2.インスリン刺激によるTSC2リン酸化はアミノ酸除去時に低下する
(A)アミノ酸除去1時間処理した細胞に、アミノ酸及びインスリン刺激(AA(+)Ins(+))、またはアミノ酸 (AA(+)Ins(–))あるいはインスリン(AA(–)Ins(+))のみ刺激を加えた際のAKT、TSC2、S6K、S6のリン酸化レベルの経時的変化をモニターしました。
(B)AA(–)Ins(+)時におけるTSC2リン酸化の低下(lane 5)は、PP2A阻害剤オカダ酸(OA)を添加することよりAA(+)Ins(+)時に観察されるTSC2のリン酸化レベル(lane 6,7)まで回復しました。
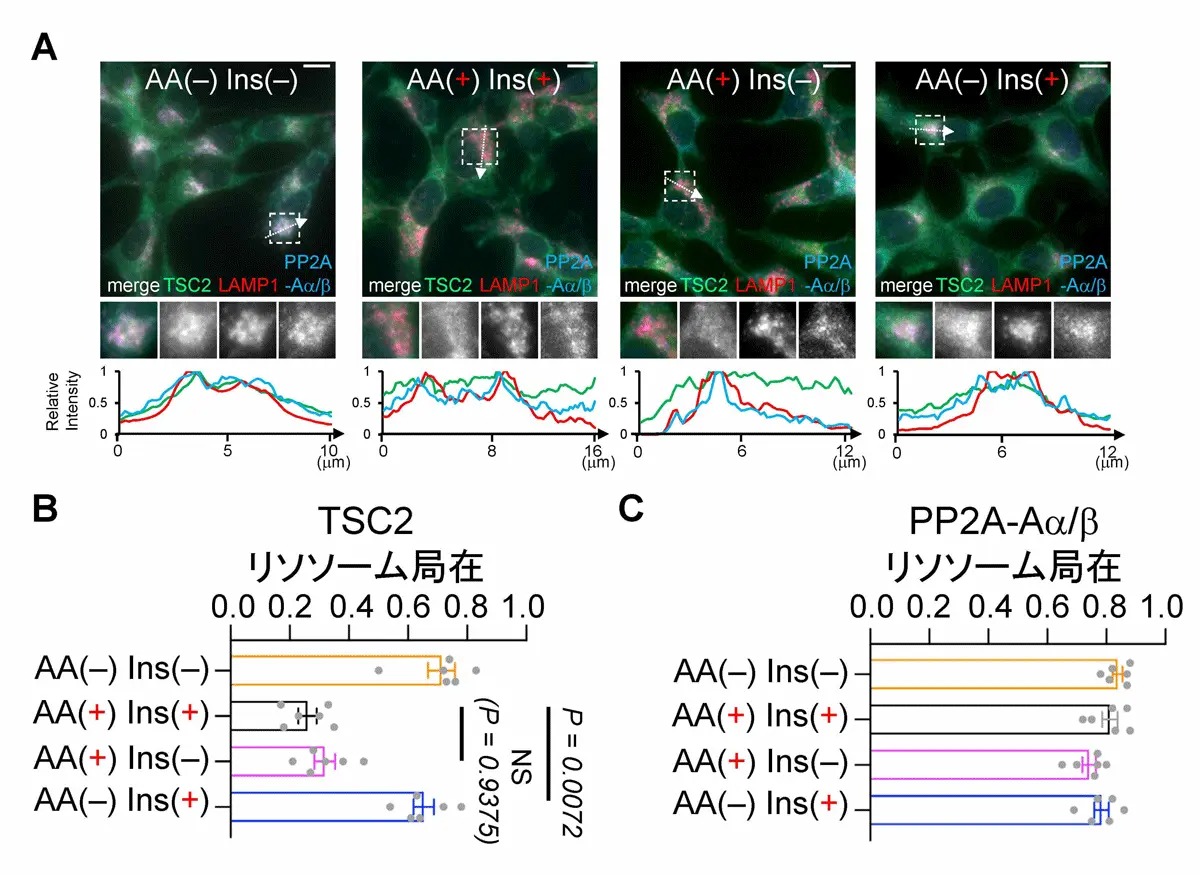
図3.TSC2はアミノ酸除去時(AA(–))にリソソーム局在が増加するが、PP2Aはアミノ酸やインスリン濃度に関係なく常にリソソーム局在を示す
(A)AA(–)Ins(–)、AA(+)Ins(+)、AA(+)Ins(–)、AA(–)Ins(+)条件におけるTSC2、PP2A-Aa/b、LAMP1(リソソームマーカー)の細胞内局在を顕微鏡撮影しました。
(B)TSC2はアミノ酸除去時(AA(–))にリソソーム局在を示しました。
(C)PP2Aはアミノ酸やインスリン濃度に関係なく常にリソソーム局在を示しました。
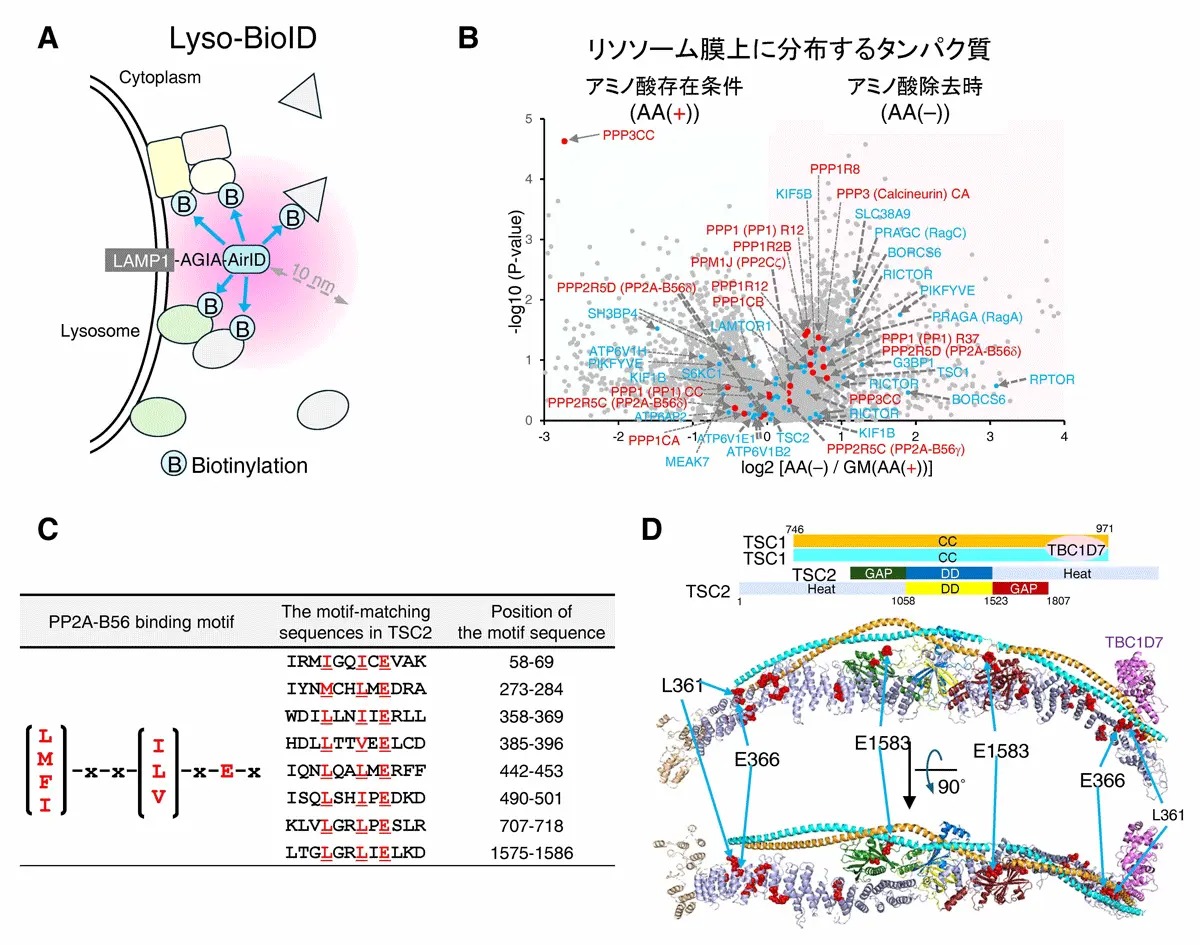
図4.PP2Aをはじめ様々な脱リン酸化酵素はリソソーム膜上に分布する
(A)近接ビオチン酵素AirIDを用いたリソソーム膜上タンパク質の網羅的ビオチン標識(Lyso-BioID)。
(B)アミノ酸存在条件(AA(+))及びアミノ酸除去時(AA(–))におけるリソソーム膜上に分布するタンパク質の質量分析(Lyso-BioID)結果。
(C)TSC2分子内に8ヶ所のPP2A-B56結合領域([L/M/F/I]-x-x-[I/L/V]-x-E-x)が存在します。
(D)PP2A-B56結合領域([L/M/F/I]-x-x-[I/L/V]-x-E-x)に、結節性硬化症でL361P変異、大腸がん、卵巣がんでE366K変異、皮膚がんでE1583K変異がそれぞれ生じていることが明らかになりました。
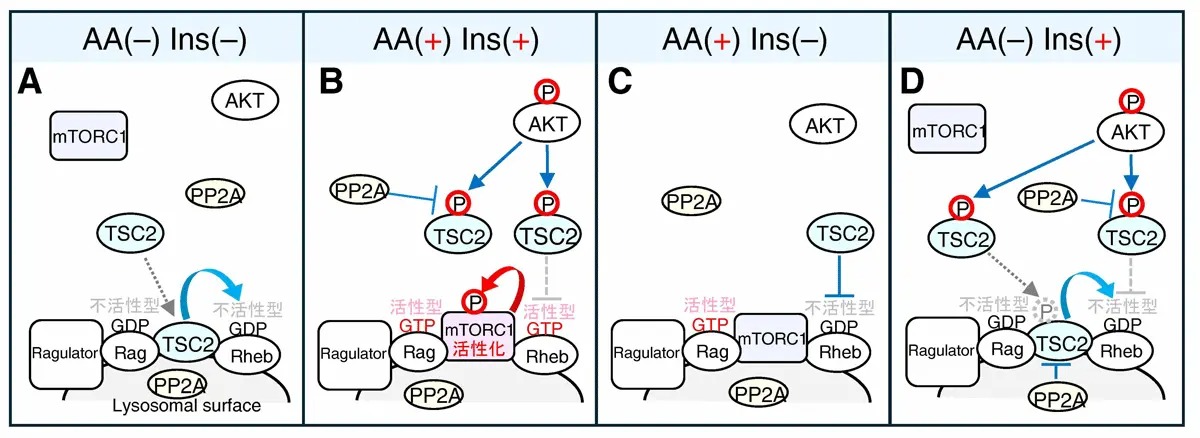
図5.リソソーム膜上でのPP2Aを介したTSC2脱リン酸化がmTORC1活性を制御する
(A)mTORC1はアミノ酸(AA)やインスリン(Ins)が存在しない場合(AA(–)Ins(–))には不活性状態にあります。
(B)アミノ酸(AA)感知経路(Ragulator-Rag)及びインスリン(Ins)感知経路(AKT-TSC1/2-Rheb)が同時に活性化された場合(AA(+)Ins(+))、Ragulator-RagGTPによってmTORC1はリソソーム膜上に移行して、RhebGTPによって活性化されます。
(C)アミノ酸経路のみ活性化した場合(AA(+)Ins(–))には、Ragulator-RagGTPによるmTORC1のリソソーム移行は誘導されますが、Rhebが不活性状態のためmTORC1の活性化は起こりません。
(D)インスリン経路のみ活性化した場合(AA(–)Ins(+))には、細胞質でAKTによってTSC2は(抑制性)リン酸化を受けますが、リソソーム膜上に移行してPP2Aによって脱リン酸化されます。脱リン酸化によりTSC2のGAP活性(ブレーキ機能)が回復するためmTORC1は不活性化状態が保たれます。
学術的・臨床的意義と今後の展望
本研究により、低アミノ酸レベルではPP2A-TSC2によってmTORC1の活性を抑制されること、さらに、PP2A-TSC2によるmTORC1活性制御の破綻は結節性硬化症(指定難病158)、あるいは、がんの発症を惹起することを明らかにしました。
また、多くのがん細胞では、遺伝子変異によりAKT、PI3Kが、恒常的に活性化しています。こうした、がん細胞ではインスリン刺激がない状態でも、インスリン感知(AKT-TSC1/2-Rheb)経路が常に活性化され続けるため、その下流のmTORC1活性も、恒常的に高い状態が維持されて、がんの進展を招きます。今回の研究成果を踏まえると、これらのがん細胞には、アミノ酸除去療法などの新たながん治療が有効であると考えられます。
謝辞
本研究は以下の研究助成を受けて行われました。
- 日本学術振興会(JSPS)科研費(21K06147、24K11046、20K06623、23K27635、19H03504)
- 日本学術振興会(JSPS)研究拠点形成事業(A. 先端拠点形成型)
- 文部科学省 新学術領域研究 「数理解析に基づく伝達システムの統合的理解」(16H06576、19H04962)
- 科学技術振興機構(JST)戦略的創造研究推進事業CREST(JPMJCR2022)
- 日本医療研究開発機構(AMED)創薬等先端技術支援基盤プラットフォーム BINDS (22ama121010j0001、23ama121010j0002)
- 武田科学振興財団 医学系研究助成
- 持田記念医学薬学振興財団 研究助成
- 高松宮妃癌研究基金 研究助成金
- 用語説明
[用語1] mTORC1(mammalian target of Rapamycin, complex 1):種を超えて高度に保存されるmTORキナーゼ(リン酸化酵素)とRAPTOR、mLST8などの制御分子から構成されるタンパク質複合体で、 Ragulator-RagA/C によってリソソーム膜表面に移行した後にRheb GTPaseによって活性化されます。
[用語2] リソソーム:細胞内小器官(オルガネラ)の1つで、その内部には消化酵素群を内包するのでタンパク質分解の場となっています。また近年はリソソーム膜表面上でmTORC1の活性化が起こるなど細胞内シグナル伝達の場として注目されています。
[用語3] TSC1/2(Tuberous Sclerosis Complex 1/2):結節性硬化症(TSC)は全身の組織で過誤腫を生じる病気(指定難病158)です。その原因遺伝子としてTSC1(9番染色体)とTSC2(16番染色体)が発見されました。TSC1とTSC2は複合体を形成してGTPase-activating protein(GAP)として機能して、mTORC1の活性化を担うRheb GTPaseの活性を抑制します。
[用語4]
iMPAQT:(in vitro proteome-assisted multiple reaction monitoring for protein absolute quantification)
内部標準タンパク質を用いて細胞内のタンパク質濃度を質量分析により網羅的に解析する手法。
[用語5]
Lyso-BioID:(proximity-dependent biotin identification for lysosomal proteins)
愛媛大学プロテオサイエンスセンターで開発されたビオチン標識酵素AirIDを活用して、リソソーム膜上に分布するタンパク質を網羅的にビオチン標識して質量分析により同定する手法。
- 論文情報
| 掲載誌: | Life Science Alliance |
|---|---|
| タイトル: |
Amino acid–dependent TSC2 dephosphorylation by lysosome–PP2A regulates mTORC1 signaling transduction
(和訳)リソソームPP2AによるTSC2のアミノ酸濃度に依存した脱リン酸化がmTORC1栄養シグナル伝達を制御する |
| 著者: |
Takanori Nakamura*, Shigeyuki Nada, Masaki Matsumoto, Nuha Loling Othman, Hidetaka Kosako, Kazuki Ikeda, Naohiko Koshikawa, Junya Masumoto, Tatsuya Sawasaki, Mutsuhiro Takekawa, Takashi Suzuki*, and Masato Okada*
*共責任著者:中村 貴紀、 鈴木 貴、 岡田 雅人 |
| DOI: |
10.26508/lsa.202503206 |
関連リンク
おすすめ記事
-
2026.07.30
-
2026.07.29
-

2026.07.28
-

2026.07.24
-

2026.07.23
![]()