生命理工学系 News
【創発的研究支援事業紹介】No.1 城所聡 助教
生命理工学院の教員が研究代表を務める研究課題が2022年度の創発的研究支援事業に採択されました。創発的研究支援事業は、特定の課題や短期目標を設定せず、多様性と融合によって破壊的イノベーションにつながるシーズの創出を目指す「創発的研究」を推進するため、既存の枠組みにとらわれない自由で挑戦的・融合的な多様な研究を、研究者が研究に専念できる環境を確保しつつ長期的に支援する文部科学省の事業です。
採択された教員をクローズアップしてご紹介するシリーズ記事(全7回)を連載いたします。全7回のうち、第1回目は“植物の温度ストレス感知機構の解明と応用”の研究課題の研究代表を務める城所聡 助教です。

城所 聡 助教
| 居室 | J2-1009室 |
|---|---|
| kidokoro.s.aa@m.titech.ac.jp | |
| Tel/Fax | 045-924-5733 |
-まず、城所先生の研究テーマを聞かせて下さい。
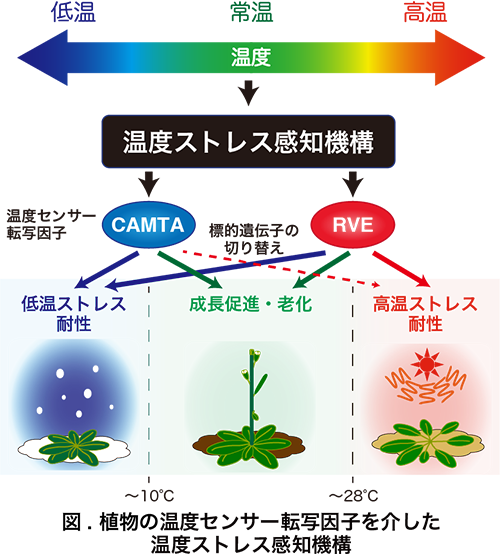
私の研究テーマは、植物における温度ストレスへの感知・応答機構の解析と応用です。植物の周囲環境は絶えず変化しており、成長や生存に多大な影響を与えています。植物は自力で動くことができないため、周囲環境の変化を速やかに感知し、遺伝子発現を大きく変化させることで環境に適応しています。生存を脅かす極度の環境変化(環境ストレス)に晒された時には、植物はストレスを耐え凌ぐための機能タンパク質をコードする数多くの遺伝子の発現を誘導します。私は、特に気温の低下による低温ストレスに応答して遺伝子発現が誘導される制御の分子機構に着目して研究を行ってきました。
高等植物の低温ストレス耐性獲得や馴化においては、DREB1という転写因子が中心的な機能を果たすことが知られています。そしてDREB1遺伝子の発現は、植物が低温ストレスを感知すると急速に誘導されます。そのため、気温の低下に応答したDREB1の遺伝子発現誘導の制御機構を解析することによって、低温ストレスの感知機構が解明できると考えています。
モデル植物であるシロイヌナズナを用いたこれまでの解析により、低温ストレスの初期応答における遺伝子発現誘導機構では、異なる転写因子群を介した2つのシグナル伝達経路が働くことを見出しました。それぞれの経路ではカルモジュリン結合ドメインを持つCAMTA転写因子群と概日時計で働くRVE転写因子群がDREB1を含めた遺伝子発現の急速な誘導を制御していました。これら2つの転写因子群は、低温ストレス時だけでなく通常生育(常温)時と高温ストレス時にもそれぞれ異なる標的遺伝子の発現を制御しており、成長・老化や高温ストレス耐性獲得に機能しています。つまりCAMTAやRVE転写因子群は、気温変化に応じて標的遺伝子を切り替える温度センサー転写因子であり、温度変化を感知して標的遺伝子を切り替える分子機構を持つと考えられます。現在はこれら温度センサー転写因子の活性制御機構に着目した研究を進めています。その結果、RVE転写因子群は常温には主に細胞質に局在しており、核には少量しか存在しませんが、低温・高温ストレス時には核へ集積すること、また低温ストレス時と高温ストレス時とでは異なるコアクチベーターが機能することを明らかにしました。
-この研究を始めたきっかけを聞かせて下さい。
私が東京大学農学部で初めて研究室に所属した時に行った研究テーマが、DREB1プロモーターから遺伝子発現を制御する領域の同定と、酵母ワンハイブリッド法によって制御領域に結合する転写因子候補として得られたタンパク質の機能解析でした。低温条件下ではレポータータンパク質の発現や活性が著しく損なわれてしまうことから、タンパク質を使ったレポーターアッセイを行うことが難しく、研究は非常に苦労しました。しかし、その分様々な実験手法を試したり、たくさんの方からアドバイスをいただいたりすることができましたので、とてもためになる経験になり研究の道を志すきっかけになったと思います。
-今回の創発的研究支援事業で取り組まれる具体的な研究内容を
聞かせて下さい。
今回の研究計画では、まず植物のCAMTAやRVE転写因子群が常温、低温、高温によって標的遺伝子を切り替えるため分子機構を明らかにするために、これら転写因子の翻訳後制御機構を解析します。RVE転写因子群を用いた解析から、温度変化による標的遺伝子の切り替えが、温度ストレスに応答した核への集積やコアクチベーターの選択といった複数段階の翻訳後制御によって行われていると考えています。そこで、温度センサー転写因子群やコアクチベーターなどの相互作用因子や翻訳後修飾といった翻訳後制御を網羅的に同定して、それら制御の機能を解析します。また、これまではシロイヌナズナを用いた解析を行ってきましたが、CAMTAやRVE転写因子群などは作物種を含む高等植物で広く保存されています。トマトなどの作物種における温度センサー転写因子の機能や翻訳後制御を解析することで、植物種ごとの温度ストレス感知機構の相違を明らかにします。
また、解明した分子機構をゲノム編集技術によって改変することで、成長と環境ストレス耐性とを両立した作物の作出技術の開発を行います。植物において成長とストレス耐性はトレードオフの関係にあります。そのため、ストレス耐性獲得に機能する遺伝子を恒常的に発現させた植物では矮化や花成遅延が起きてしまうことがストレス耐性作物の開発において大きな課題となっています。成長とストレス耐性の切り替えスイッチとなる温度ストレス感知機構をゲノム編集技術によって改変することで、成長と環境ストレス耐性とを両立した作物の作出を目指します。
-今後の目標を聞かせて下さい。
今後の目標は、植物の温度ストレス感知機構を明らかにすることです。ヒトを含む多くの生物では、カルシウムイオンなどの陽イオンを細胞内に流入させるTRPチャネルファミリーが温度センサーとして働くことが知られています。カプサイシン受容体TRPV1が熱を感知することを最初に同定した研究者は2021年にノーベル医学・生理学賞を受賞しました。またTRPチャネルファミリーを創薬のターゲットとした応用研究も世界中で行われています。一方で、植物にはTRPチャネルの相同タンパク質が保存されていないことから、植物独自の温度感知機構が存在すると考えられています。植物の生育環境は多様であり、生育至適温度やストレスとして感知する温度は種によって異なります。創発的研究支援事業での研究を通して、植物が持つ温度ストレス感知の分子機構を明らかにすることにより、植物が周囲温度環境に適応する基本原理を理解したいと考えています。また解明した分子機構を元に成長やストレス耐性などをコントロールする様々な技術の開発を行うことで、様々な気候変動に対応できる農作物の開発に繋げたいと考えています。
おすすめ記事
-
2026.07.30
-
2026.07.29
-

2026.07.28
-

2026.07.24
-

2026.07.23
![]()